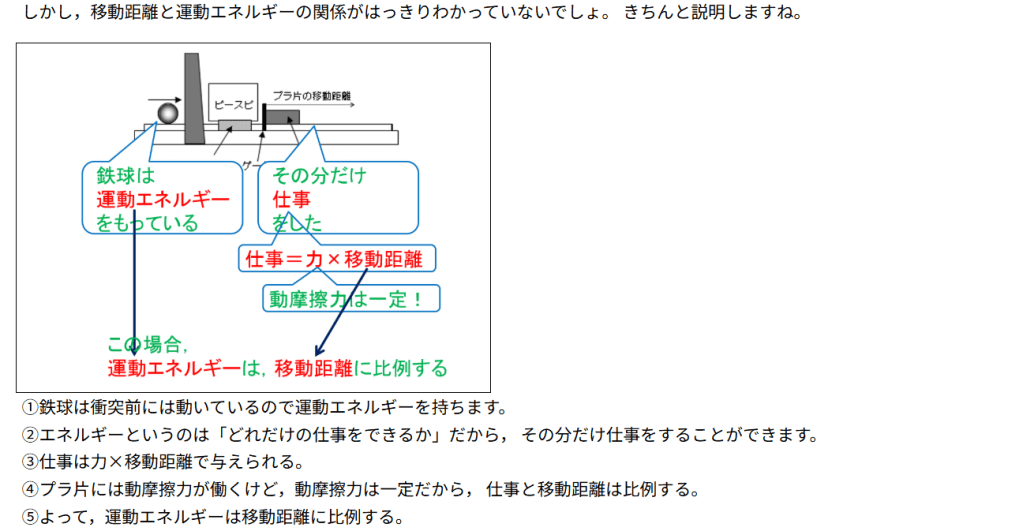
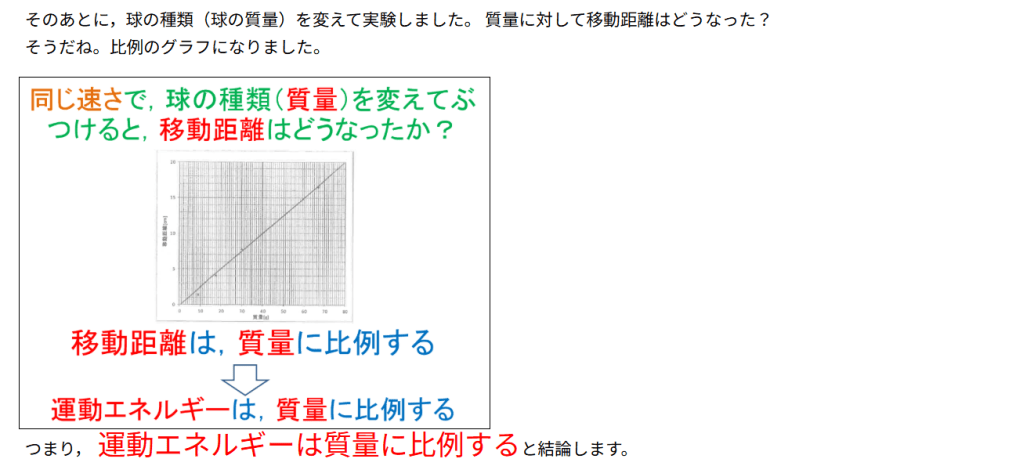
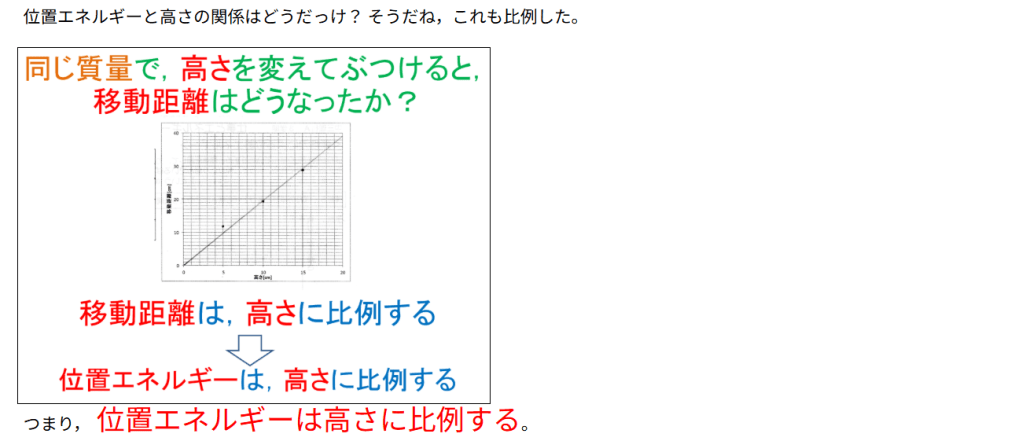
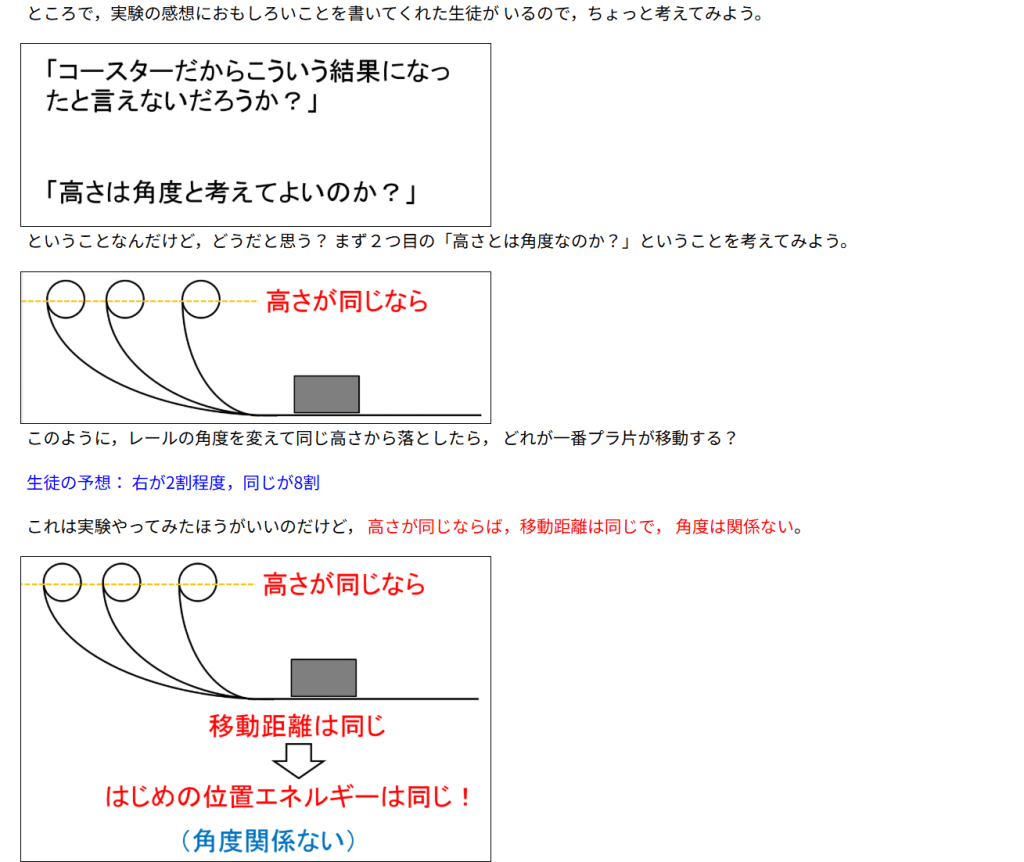
カテゴリー: サイエンス
-
-
『AI による概要
図解!エントロピー増大の法則とは?自発変化の方向を示す熱力学 …
【東大院生が解説】エントロピーって結局何?~やたら抽象的 …
図解!エントロピー増大の法則とは?自発変化の方向を示す熱力学 …
図解!エントロピー増大の法則とは?自発変化の方向を示す熱力学 …
図解!エントロピー増大の法則とは?自発変化の方向を示す熱力学 …
Untitled
すべて表示エントロピーが運動の「質」まで把握できるようになった、とは、単なる「乱雑さ」の度合いだけでなく、熱や運動の持つエネルギーがどれだけ利用可能な仕事(エネルギーの質)に変換できるかを評価できるようになったことを指します。
エントロピー増大の法則により、系が自然に放置されるとエネルギーは質が低い状態(エントロピーが高い状態)へと向かう傾向にあると理解され、この性質を通じてエネルギーの変換効率や状態の変化を、より本質的に捉えられるようになったのです。
具体的な意味合い
古典的なエントロピーから「エネルギーの質」へ
当初、エントロピーは熱力学で「無秩序さの度合い」として定義されました。
例えば、部屋が散らかっている状態(高エントロピー)と片付いている状態(低エントロピー)のようなイメージです。
しかし、この概念は、熱エネルギーがどれだけ有用な仕事に変換できるか、つまりエネルギーの「質」を評価するのにも応用できるようになりました。
エネルギーの質とエントロピーの関係
高エントロピー:のエネルギーは、質の低いエネルギーです。
たとえば、熱が広範囲に拡散した状態は熱運動が激しく(高温で)乱雑なためエントロピーが高く、仕事への変換効率も低くなります。
逆に、低エントロピーのエネルギーは、質の高いエネルギーであり、構造化されたエネルギーで、より有用な仕事に変換することが可能です。
「エントロピー増大の法則」による示唆
エントロピー増大の法則は、外部からの仕事が加えられない限り、系は常にエントロピーが増大する方向へ進むというものです。
これは、エネルギーが自然に質が低く、乱雑な状態へと移行していく傾向があることを示しています。
つまり、運動の「質」は自然に低下していく、と捉えることができるのです。
このように、エントロピーの概念は、単に「乱雑さ」を指すだけでなく、エネルギーの持つ質や、エネルギーの変換効率といったより深い側面を理解するための重要な指標となったのです。
1分子1分子の乱雑さからエントロピー変化を定量する
2024/05/31 — 発表内容 蒸機機関の効率化に端を発した熱力学研究成果の金字塔が、クラウジウスが1854年に提案したエ…東京大学 大学院理学系研究科・理学部
熱運動の原理と産業・医療への応用 – 下西技研工業株式会社
原子や分子の熱運動が激しいほど、エントロピーは高くなります。 逆に、温度が低くなるほど、熱運動は小さくなりエントロピーは…下西技研工業
レポート ・ エントロピー増大の法則
エントロピーという言葉を使って表現すれば、すべての事物は、「それを自然のままにほっておくと、そのエントロピーは常に増大し…
ワシモ(WaShimo)のホームページすべて表示
AI モードでさらに詳しくAI の回答には間違いが含まれている場合があります。 詳細 』
-
『物理学・化学
「エントロピー」という概念がよくわかりません。部屋は汚くなるが、キレイにはならない、みたいな例えをたまに聞きますが。。。良ければこの概念を理解するために有益そうなことを教えて頂きたいです。』https://mond.how/ja/topics/25cvmio3xol00zd/t242v2yde410hdy
『白石直人
匿名フォロー
東京大学総合文化研究科広域科学専攻相関基礎科学系准教授。専門は理論物理、特に非平衡統計力学。 アウトリーチ活動にも関心あり。過去には仮面ライダービルドの物理学アドバイザーも務める。 人文・社会科学系含めて読書は幅広く行っており、別冊文藝春秋にてブックガイド記事を連載中。「エントロピー」が何であるのか理解しづらいのは、エントロピーが複数の分野で異なる対象に対して異なる方法で定義されているからです。
勿論異なる定義の量が同じ「エントロピー」という名前で呼ばれているのは、それらに似ている側面が少なくないからですが、しかし基本的設定や定義が異なることは意識しておくとよいでしょう。
熱力学は、「マクロな平衡状態の系の遷移」を扱う分野です。
平衡状態というのは、大雑把に言えば、完全に緩和が終わった後の、流れもなく一様な状態のことです。例えば一様な水とか気体とかはその例です。
熱力学における「エントロピー」は、断熱操作で状態が移り変われるかを特徴づける量として定義できます。
断熱操作というのは、イメージとしては「絶対熱が逃げない魔法瓶」の中にあるような状況です。外とのエネルギーのやり取りは、ピストンを押したり引いたりすることによってのみ可能です。
このような断熱操作で、ある状態Xが別の状態Yに移り変われるには、XのエントロピーS(X)がYのエントロピーS(Y)よりも小さい必要があります。
言い換えれば、エントロピーは減ることがなく、常に増えていくということです。
これは「エントロピー増大則」と呼ばれています。熱力学のエントロピーは、変化の不可逆性を特徴づけている、ともしばしばいわれます。
平衡統計力学は、「対象とする物体のミクロな知識から、マクロな性質(エントロピーなどの量)を導く」ことを行う分野です。
平衡統計力学でもエントロピーが定義されており、これは「ボルツマンエントロピー」とも呼ばれます。
ある物体のあるマクロな平衡状態におけるボルツマンエントロピーは、対象となる物体がそのマクロな平衡状態であるような「とりうるミクロな状態の総数」の対数で定義されます。
例えばマクロな平衡状態がエネルギーE、体積V、粒子の数Nで指定されているのならば、「とりうる状態の総数」というのは、体積Vの容器中にN個の粒子を配置する方法で、エネルギーがEであるような方法の総数のことです。
とりうるミクロな状態の数が多ければ多いほど、ボルツマンエントロピーは大きくなります。
平衡統計力学のボルツマンエントロピーは、とりうる状態の広さを特徴づけている、とも言われます。
そして、ミクロな方法で定義されたボルツマンエントロピーは、マクロな状態の変化可能性を用いて定義された熱力学のエントロピーと一致します。これは「ボルツマンの原理」と呼ばれているものです。
エントロピーは、物理だけでなく情報理論でも用いられています。
情報理論における「シャノンエントロピー」は、確率的に生じる事象に対して定義されており、そこで生じる事象がどれくらい不確かなのか(予測しづらいか)を定量化しています。
何が起きるのかを予測しやすい場合(例えば、(重りを仕込んでいるので)高い確率で6の目が出るサイコロを振る)にはシャノンエントロピーは小さい値をとり、何が起きるのかを予測しづらい場合(例えば、均等なサイコロを振る)にはシャノンエントロピーは大きな値をとります。
情報理論のシャノンエントロピーは、乱雑さを特徴づけている、とも言われます。
シャノンエントロピーとボルツマンエントロピーにもつながりがあります。
あるマクロな平衡状態に対応するミクロな状態がすべて等確率で出現するとします。
すると、この状態出現に対するシャノンエントロピーは、ボルツマンエントロピーに一致することが示せます。
ただし、両者は異なる対象に対して定義されていることは意識しておく必要があります。
サイコロに対してはシャノンエントロピーは定義可能ですが、ボルツマンエントロピーは定義されていません。注意しておきたいのは、熱力学のエントロピーも、平衡統計力学のボルツマンエントロピーも、マクロな平衡状態に対して定義されている量だということです。
マクロな平衡状態ではない対象、例えばあなたの部屋、に対しては、これらの量はどちらも定義されていません。
また、シャノンエントロピーは確率的に生じる出来事に対して定義されている量です。
もしあなたの部屋の状況が、サイコロを振った際の出目のように「何か確率的に事象が起きる」とみなせるのならば、あなたの部屋のシャノンエントロピーは定義できます。
しかしそうでないならば、あなたの部屋のシャノンエントロピーという量は定義されません。
世間で用いられる「エントロピー」の少なくない場合は、これらのエントロピーがどれも定義されていない対象に対して、ただアナロジーとして用いられています。
』 -
『熱力学においてエントロピーを考える計算上のメリットは、系が自然に変化する方向や究極の状態を判断する尺度となることです。エントロピー増大の法則により、エントロピーが増加する方向への自発的な変化は計算可能になり、エントロピーが最大値に達すると平衡状態に達することがわかります。これにより、エネルギーの流れや仕事の変換効率など、現実世界の熱力学現象の予測や分析を容易にします。
具体的なメリット
自発的プロセスの予測と判断
: 自然に起こるプロセスはエントロピーが増加する方向へ進みます。エントロピーという概念を導入することで、あるプロセスが自発的に進行するかどうか、またその進行方向を定量的に判断する基準が得られます。平衡状態の判定
: 系が非平衡から平衡状態へと向かうとき、エントロピーは最大値に達します。エントロピーの増大という視点から計算することで、系が最終的にどのような平衡状態に落ち着くかを予測できます。エネルギー変換効率の限界の理解
: 熱機関の効率など、エネルギーの変換プロセスにおけるエントロピーの増大を追跡することで、実現可能な最大効率の限界を計算的に理解し、評価することができます。マクロな現象のミクロな視点からの説明
: 統計力学的な観点からエントロピーは、多数の分子の配置の「場合の数」の多さ、すなわちミクロな状態の多様性に対応します。このミクロな視点を取り入れることで、マクロな状態変化(不可逆過程)がなぜ起こるのかを計算的に説明できるようになります。エントロピー – Wikipedia
エントロピー(英: entropy)は、熱力学や統計力学、情報理論などにおいて定義される示量性の状態量のひとつである。 エントロピーはエネルギーを温度で割った次元を持ち、国際単位系における単位はジュール毎ケルビン(記号: J/K)である。 エントロピーと同じ次元を持つ量として熱容量がある。
お気に入りアイコン(ファビコン)
Wikipedia【東大院生が解説】エントロピーって結局何?~やたら抽象的な概念を …
Jun 22, 2025 — この統計力学的な解釈は、熱力学の「エントロピー増大の法則」にミクロな視点からの裏付けを与えました。 つまり、自然は、より「確率の高い」状態、すなわち「場合の数が多い」状態へと自発的に移り変わっていく。 これが、エントロピーが増大する本当の理由だったのです。
お気に入りアイコン(ファビコン)
note熱力学の基礎入門~数学的な基本原理から熱力学を学ぶ~|マスログ
Oct 3, 2021 — が最大になるとき,その場合に限り複合系は平衡状態になります. その時の複合系のエントロピーは\hat{S} の最大値になります.
お気に入りアイコン(ファビコン)
和から株式会社すべて表示
AI の回答には間違いが含まれている場合があります。 詳細 』 -
『熱力学におけるエントロピーは、「システムの乱雑さ(無秩序度)の度合い」を示す物理量であり、エントロピーが増加する方向に自然現象が進行することを示す「熱力学第二法則」の中心的な概念です。
具体的には、熱力学系が吸収した熱量をそのときの絶対温度で割った値として定義され、不可逆なプロセス、つまり元に戻れない自然現象(例:水に垂らしたインクが広がる)でエントロピーは増大します。
エントロピーの主な特徴
乱雑さの指標:
秩序がある状態(例:氷の結晶)はエントロピーが低く、乱雑な状態(例:水になって動き回る分子)はエントロピーが高いとされます。不可逆性の表現:
熱は高温から低温へ流れるように、自然界の多くの変化はエントロピーが増加する方向に進みます。例えば、広がったインクは自然に元の1滴に戻ることはありません。計算方法:
熱力学的なエントロピー変化は、可逆的な過程で系に吸収された熱量(δQ)をその時点の絶対温度(T)で割った値(δQ/T)を積分することで得られます。熱力学第二法則:
孤立した系では、エントロピーは常に増大するか、変化が全くない場合でも一定に保たれます。これは自然に起こりうるすべての現象がこの法則に従うことを意味します。例による理解
インクが広がる現象:
水にインクを1滴落とすと、インク分子は水の中で広がり、全体に拡散します。この拡散した状態は、もとの1滴の状態よりも分子が乱雑であるため、エントロピーが増大した状態と言えます。氷が溶ける現象:
氷が溶けて水になると、氷の結晶構造に固定されていた水分子が自由に動き回るようになります。これにより分子の動きがより自由で乱雑になるため、エントロピーが増加します。エントロピー – Wikipedia
エントロピーはエネルギーを温度で割った次元を持ち、国際単位系における単位はジュール毎ケルビン(記号: J/K)である。 …Wikipedia
「エントロピー」って何? 複雑な定義を身近な事例でわかり …
2024/04/23 — 熱力学における重要な概念 エントロピーは、物理学の一分野である「熱力学」に登場する概念です。 熱力学…HugKum
図解!エントロピー増大の法則とは?自発変化の方向を示す熱力学の …
2021/04/28 — 1. エントロピーとは?(簡単な説明) エントロピーとは簡単にいうと「無秩序な状態の度合い(=乱雑さ…jp.dreamscope.me
すべて表示
AI の回答には間違いが含まれている場合があります。 詳細 』 -
アデノシン三リン酸(wiki)
https://ja.wikipedia.org/wiki/%E3%82%A2%E3%83%87%E3%83%8E%E3%82%B7%E3%83%B3%E4%B8%89%E3%83%AA%E3%83%B3%E9%85%B8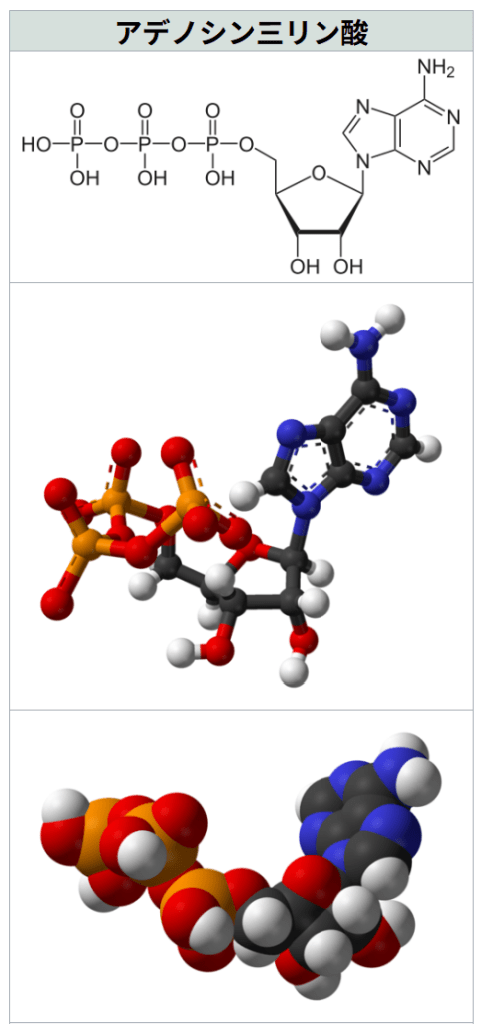

『出典: フリー百科事典『ウィキペディア(Wikipedia)』
アデノシン三リン酸{{{画像alt1}}}
{{{画像alt2}}}
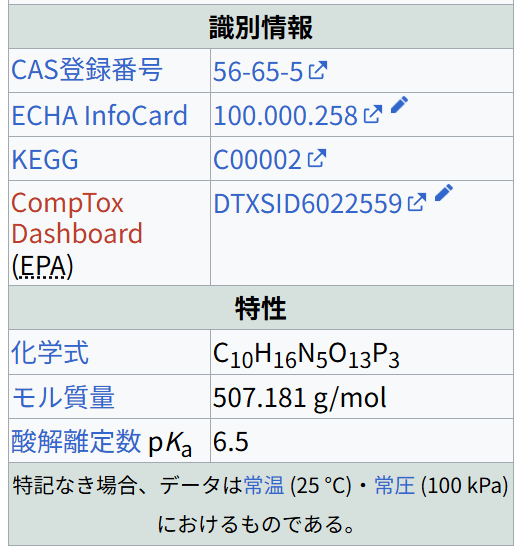
IUPAC名[表示]識別情報
CAS登録番号
56-65-5
ECHA InfoCard 100.000.258 ウィキデータを編集
KEGG
C00002
CompTox Dashboard (EPA)
DTXSID6022559 ウィキデータを編集
特性
化学式 C10H16N5O13P3
モル質量 507.181 g/mol
酸解離定数 pKa 6.5
特記なき場合、データは常温 (25 °C)・常圧 (100 kPa) におけるものである。アデノシン三リン酸(アデノシンさんリンさん、英: adenosine triphosphate)とは、アデノシンのリボースに3分子のリン酸が付き、2個の高エネルギーリン酸結合を持つヌクレオチドである。
リボースの5位の炭素に、リン酸が結合しているため、アデノシン 5′-三リン酸などとも書かれる。
しばしば「adenosine triphosphate」から取ったアルファベットを並べて「ATP(エー・ティー・ピー)」と呼称される。本稿では以後、ATPと略記する。
所在
ATPは真核生物や真正細菌など、既知の地球生物の全ての細胞が利用している解糖系でも産生される物質であるため、地球上の生物の体内に広く分布する。
生体内では、リン酸1分子、または、リン酸2分子が離れたり結合したりする事で、エネルギーの放出・貯蔵を行う[1]。
なお例えば、糖に限らず、真核生物が脂肪酸やアミノ酸などをエネルギーとして利用する際も、例えば、一部はGTPに変換されて、そのままGTPが別の用途に用いられる場合など例外はあるものの、主にATPに変換してからエネルギーとして利用し、色々な用途に活用している。
これらの理由のため、既知の地球生物の各細胞には普遍的にATPが存在する。
なお、しばしば地球生物の細胞は、ATPを経由して物質のエネルギーを利用しているため、ATPは「生体のエネルギー通貨」とも形容される。
構造とエネルギー
プリン塩基であるアデニンに、単糖のリボースがN-グリコシド結合により結合したアデノシンを基本構造として、リボースの 5′-ヒドロキシ基にリン酸エステル結合によりリン酸基が結合し、さらにリン酸が2分子連続して無水結合で結合した構造である。
この、リン酸基同士の結合(リン酸無水結合)は、エネルギー的に不安定であり、このリン酸基の加水分解による切断反応や、他の分子にリン酸基を転移させる反応(切断した両リン酸基の端に、反応により新たに生成する、より安定な化学結合の生成に伴って)で、エネルギーを放出する。
ATPのリン酸基の加水分解や転位反応は、正味の自由エネルギーの減少を伴うエネルギー放出反応であり、あたかもATPのリン酸基同士の結合の切断が生体内の化学反応の実質的な推進力であるかのように見えるため、この意味において、この結合は「高エネルギーリン酸結合」と呼ばれており、これはリン原子が3つ繋がった状態である[2]。
エネルギーの収支式を以下に示す(ΔG°’(標準自由エネルギー変化))。
ATP + H2O → ADP(アデノシン二リン酸) + Pi(リン酸)
ΔG°’ = −30.5 kJ/mol (−7.3 kcal/mol)
ATP + H2O → AMP(アデノシン一リン酸、アデニル酸) + PPi(ピロリン酸)ΔG°’ = −45.6 kJ/mol (−10.9 kcal/mol)
この標準自由エネルギー変化は、一般的なリン酸エステル化合物のリン酸エステル結合の加水分解の標準自由エネルギー変化(ΔG°’ = −3〜4 kcal/mol)などに比べ非常に大きいので、このようなリン酸エステル化合物が、ATPからのリン酸基の転移により生成する反応の標準自由エネルギー変化は、全体として負の値であり、この反応はATPからリン酸エステル化合物へのリン酸転移の方向に自発的に進む。
さらに細胞内では、ATP濃度はADPの10倍程高く、リン酸濃度も標準状態 (1.0 M) より、はるかに低い (1〜10 mM程度) ため、細胞内の環境ではATPの高エネルギーリン酸結合の加水分解に伴って実際に放出されるエネルギー(自由エネルギー変化 ΔG)は、より大きく、−10〜11 kcal/mol に達する。
生合成
ATPは主にATP合成酵素において酸化的リン酸化、光リン酸化によって生じる。
ADP + Pi → ATP
また、解糖系やクエン酸回路などでもATPは合成される。
→好気呼吸によるATPの収支式については「好気呼吸」を参照
GTP(グアノシン三リン酸)については、以下の反応式でATPと相互変換する。
GTP + ADP ⇔ GDP + ATP (ΔG°’ 〜0)
また、細胞内では、アデニル酸キナーゼの働きにより、ATP, ADP, AMPが次の反応による平衡混合物として存在し、ATPはADPからも一部再生される。
2 ADP ⇔ ATP + AMP (ΔG°’ 〜0)
ATPの役割
ATPはエネルギーを要する生物体の反応素過程には必ず使用されている。
例えば、哺乳類の骨格筋100 gあたりに、ATPは0.4 g程度存在する。反応・役割の例については、以下の物などが挙げられる。
解糖系 – グルコースのリン酸化など。
筋収縮 – アクチン・ミオシンの収縮。
能動輸送 – イオンポンプなど。
生合成 – 糖新生、還元的クエン酸回路、尿素回路など。
タンパク質の形状正常化 – タンパク質の安定化に関わる分子シャペロンもATPを利用する。
原料 – アデニル酸サイクラーゼによって、ATPはcAMPに変換される。また、RNA合成の前駆体などでもある。発光タンパク質 – ルシフェラーゼなど。
発電 – 電気ウナギに見られる筋肉性発電装置。
発熱 – 反応の余剰エネルギーなど。
なお、リン酸基の付加はリン酸基転移酵素(キナーゼ)によって行われる。
用途
有効成分としてATP-2Naを配合した内服薬
ATPは、医薬品としても利用されている。
日本では2011年現在、調節性眼精疲労の症状改善、消化管機能低下が起きている者の慢性胃炎の症状改善、心不全の症状改善、頭部外傷後遺症の症状改善に用いられる[3]。
この他、2017年現在、日本ではATPの顆粒製剤のみは、メニエール病や内耳障害を原因とするめまいの改善にも用いられる[4]。
なお、消化管機能低下が起きている者の慢性胃炎については軽症患者の自覚症状の改善に有効だったとされている[5]。
歴史
1929年 – Fiske、Subbarowら、そしてLoehmannによって独自に、不安定なリン酸結合を持つヌクレオチドとして発見された。
当初、ATPはエネルギー通貨ではなく、リン酸供与体の一部として認識されていた。
1931年 – Loehmann、Meyerhofによって解糖系にATPが用いられる事が明らかになった。
1939年 – Engelhardtらによって、筋収縮のタンパク質であるミオシンが、ATPを加水分解する活性を有する事が明らかになった。
同年、フリッツ・アルベルト・リップマンによってATPは代謝に中心的な役割を果たしている事が提唱された。
1941年 – セント=ジェルジ・アルベルトによってミオシンが、ATPによって収縮する事が明らかになった。
1942年 – セント=ジェルジによってアクチン、ミオシン、ATPが筋収縮の基本的な構成単位である事が明らかになった。
これらのハンガリー学派の筋収縮に関する一連の研究が「ATPは生体のエネルギー通貨」であるという認識を構築していった。
また、ATPが能動輸送に関係することが1957年、イェンス・スコウらによって明らかにされ(Na+, K+-ATPaseの発見)、ATP利用系のフォーマットが現在に至るまで構築されている。
ATP合成系の歴史については、以下の通りである。
1951年 – Lehningerによって呼吸鎖複合体の電子伝達およびATPの合成は共役しているという「酸化的リン酸化」が提唱された。
1961年 – Mitchellによってプロトンの電気化学ポテンシャルがATPの合成に寄与していると言う「化学浸透圧仮説」が提唱された。
1963年 – Avronによって葉緑体のチラコイド膜上に球状突起が見出され、この構造体がATP合成に関係した酵素であると推定された。
1966年 – Jagendorfらによって葉緑体でのpHジャンプによるATP合成系のモデルが提唱された。
1975年 – RackerとStoeckeniusによって、脂質二重層を用いたATP合成酵素およびバクテリオロドプシンの実験によって、ATP合成が電気化学的ポテンシャルによって行われる事を明らかにした。
1978年 – 化学浸透圧説を唱えたMitchellがノーベル化学賞を受賞した。
1981年 – BoyerがATP合成酵素の「回転触媒仮説」を提唱した。
1994年 – WalkerらによってウシATP合成酵素のF1サブユニットのX線結晶構造解析が行われ、その立体構造が明らかになった。
1997年 – Boyer、WalkerらがATP合成酵素の反応素過程を解明した業績で、ノーベル化学賞を受賞した。
2008年 – 岡山大学の森山芳則教授らの研究グループがATPのトランスポーターを特定し、3月25日『米国科学アカデミー紀要 (PNAS)』電子版に掲載・発表された論文において、これを「小胞型ヌクレオチド・トランスポーター (vesicular nucleotide transporter, VNUT)」と命名した[6][7]。
脚注・参考文献
^ デジタル大辞泉【アデノシン三リン酸】(アデノシンさんりんさん)
^ ただし、結合自体がエネルギーを持つわけではない:この化学結合の切断は、吸エネルギー反応である。
^ ATP腸溶錠(p.1)
^ ATP腸溶錠・ATP顆粒剤(p.11)
^ ATP腸溶錠・ATP顆粒剤(p.15)
^ Sawada, K.; Echigo, N.; Juge, N.; Miyaji, T.; Otsuka, M.; Omote, H.; Yamamoto, A.; and Yoshinori Moriyama (April 15, 2008) “Identification of a vesicular nucleotide transporter” Proceedings of the National Academy of Sciences of the United States of America, 2008 105: 5683-5686; doi:10.1073/pnas.0800141105
^ VNUTによって神経末端のシナプス小胞に運ばれたATPは貯蔵された後、外部に放出されて疼痛を発生させたり血管を収縮したりするため、VNUTが抑制できれば痛み・血管収縮を管理することが可能かもしれないと考えられている。
関連項目ウィキメディア・コモンズには、アデノシン三リン酸に関連するカテゴリがあります。
呼吸
解糖系
電子伝達系
ATP合成酵素(ATPシンターゼ)
アデノシン三リン酸フォスファターゼ(ATPアーゼ)
環状アデノシン一リン酸 (cAMP)
ATP測定法(ATP拭き取り検査)
表話編歴
核酸の構成要素
表話編歴
補因子
表話編歴
食品科学
表話編歴
代謝、異化、同化
表話編歴
代謝マップ
典拠管理データベース ウィキデータを編集
カテゴリ: 代謝経路生体物質プリンヌクレオチド補酵素最終更新 2025年6月20日 (金) 08:18 (日時は個人設定で未設定ならばUTC)。
テキストはクリエイティブ・コモンズ 表示-継承ライセンスのもとで利用できます。追加の条件が適用される場合があります。詳細については利用規約を参照してください。』 -
ミトコンドリア
https://ja.wikipedia.org/wiki/%E3%83%9F%E3%83%88%E3%82%B3%E3%83%B3%E3%83%89%E3%83%AA%E3%82%A2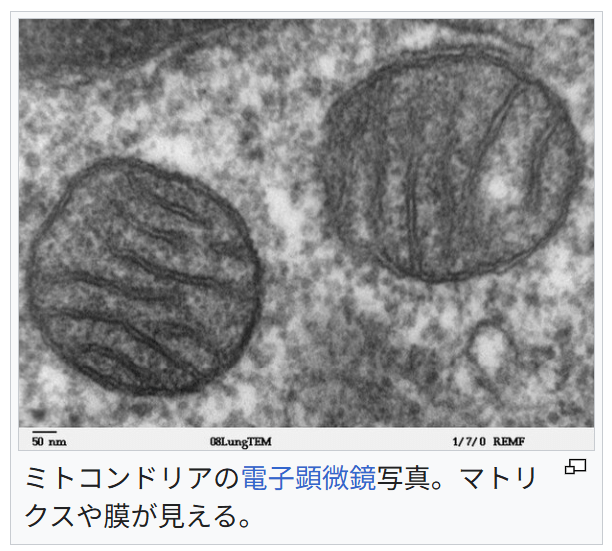

『出典: フリー百科事典『ウィキペディア(Wikipedia)』
ミトコンドリア(英語: mitochondrion、複数形: mitochondria)は、ほとんど全ての真核生物の細胞の中に存在する、細胞小器官の1つである。シトクロムcを含むため本来は赤味を帯びるがヤヌスグリーンによって青緑色に染色される。
ミトコンドリアの電子顕微鏡写真。マトリクスや膜が見える。
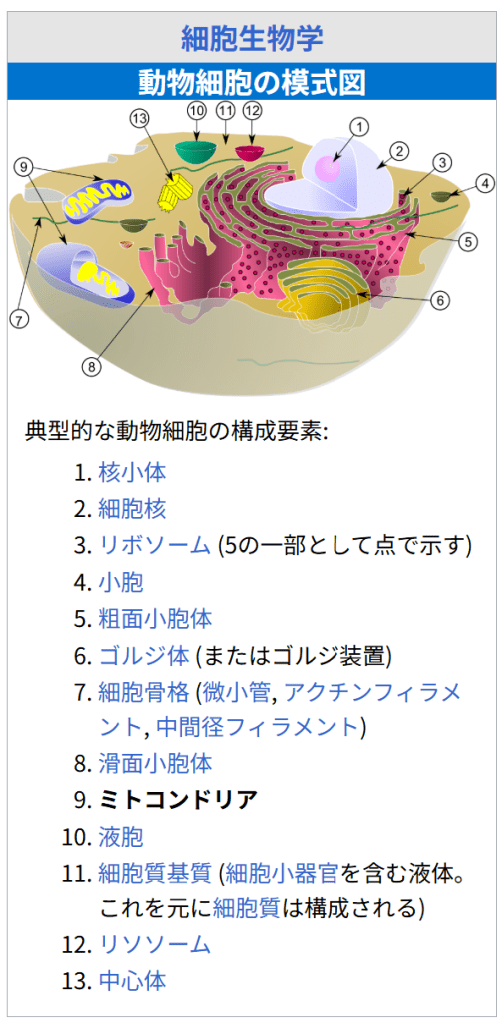
細胞生物学
動物細胞の模式図典型的な動物細胞の構成要素:
核小体
細胞核
リボソーム (5の一部として点で示す)
小胞
粗面小胞体
ゴルジ体 (またはゴルジ装置)
細胞骨格 (微小管, アクチンフィラメント, 中間径フィラメント)
滑面小胞体
ミトコンドリア
液胞
細胞質基質 (細胞小器官を含む液体。これを元に細胞質は構成される)
リソソーム
中心体ミトコンドリアは脂質二重層でできた外膜と内膜を有し、膜には様々なタンパク質が存在する。
ミトコンドリアでは、高エネルギーの電子と酸素分子を利用して、ATPを合成する。
すなわち、ミトコンドリアは真核生物における好気呼吸の場である。
また、真核生物の細胞が有する核とは別に、ミトコンドリア独自のミトコンドリアDNA(mtDNA)を内部に有し、ある程度ながら自立的にミトコンドリアは細胞内で分裂して、増殖する。
このmtDNAは、ミトコンドリア内部だけに限らず、真核生物の細胞全体の生命現象にも関与する。
さらに、細胞のアポトーシスにおいても、ミトコンドリアは重要な役割を担っている。
ヒトにおいては、肝臓、腎臓、筋肉、脳などの代謝の活発な細胞には特に多くのミトコンドリアが存在し、細胞質の約40パーセントを占めている。
全身の平均では、1細胞中に300個から400個のミトコンドリアが存在し、全身で体重の約1割を占めていると概算されている[1]。
単語の「Mitochondrion」はギリシャ語のμίτος, mitos「糸」とχονδρίον, chondrion, 「顆粒」に由来し[2]、糸粒体(しりゅうたい)と和訳される例も見られる。
構造
ミトコンドリアの構造
1.内膜 2.外膜 3.クリステ(平板状) 4.マトリクス
ミトコンドリアの直径は0.5 μm程度であるが、その形状は、生物種や細胞の置かれている条件によって多様である。
球形、円筒形、紐状、網目状など様々な形状のミトコンドリアが存在し、長さが10 μmに達する物も珍しくない。
1細胞あたりの数は、1つに維持されている細胞もあるが、多い場合では数千個のミトコンドリアが絶えず分裂と融合を繰り返している場合もある。
ミトコンドリアは外膜と呼ばれる脂質膜に囲まれており、その内側に、もう1枚、内膜と呼ばれる脂質膜を有する。
内膜に囲まれた内側をマトリクス[注釈 1]、内膜と外膜に挟まれた空間を膜間腔と呼ぶ。
なお、内膜はマトリクスに向かって陥入した、クリステ(cristae)と呼ばれる特徴的な構造を取っている。参考までに、この「cristae」とは「櫛」という意味である[3]。ミトコンドリアは照射された光を強く屈折するため、生きた細胞を位相差顕微鏡で観察すると、ミトコンドリアが明瞭に確認できる[3]。
生きた細胞を観察すると、ミトコンドリアが細胞内で、伸縮したり、屈曲したりと、動いている姿も確認できる[3]。
外膜
真核生物の細胞膜と同様に、ミトコンドリアの外膜の組成も、タンパク質とリン脂質の重量比が約1:1である。
外膜の進化的起源は真核生物の細胞内膜系だと考えられ、現在でも小胞体膜と物理的に関係しており、カルシウムシグナルの伝達や脂質の交換を行っている[4]。
外膜にはポリンと総称される膜タンパク質が大量に存在し、分子量5000以下の分子が、外膜を通過できるようなチャネルを形成している。
これより大きなタンパク質は自由に出入できず、タンパク質のペプチド配列中に、特別な移行シグナルが付与されている場合にのみ、細胞質からミトコンドリア内へと取り込まれる[5]。
膜間腔
→詳細は「ミトコンドリア膜間腔」を参照膜間腔は、ミトコンドリアの外膜と内膜に挟まれた空間である。
外膜がポリンによって低分子を自由に透過させる性質を実現しているため、通常の状態において、膜間腔のイオンや糖などの組成の多くは、ほとんど細胞質と同等である。
例外は、内膜の直近のプロトンの濃度のように、限られる。
その一方で、膜間腔におけるタンパク質の組成は、細胞質と大きく異なっており、外膜が破壊されて膜間腔に存在するタンパク質(シトクロムcなど)が細胞質へと漏れ出すと、細胞のアポトーシスが引き起こされる[6]。
内膜
内膜はマトリクスと膜間腔とを隔てており、ミトコンドリアの機能的アイデンティティを担っている。
酸化的リン酸化に関わる呼吸鎖複合体などの酵素群が、内膜には規則的に配列している。
外膜とは対照的に、基本的に内膜は不透性であり、何らかの物質を内膜を横断して輸送するためには、それぞれの物質に対して特異的な輸送体が必要である。呼吸鎖複合体は内膜を挟んで、マトリクスからプロトンを膜間腔へと汲み出して、膜間腔の側のプロトンの濃度を高め、濃度勾配が形成される。
この濃度勾配が、物質輸送やATP合成に関与している。
また、マトリクスへのタンパク質輸送装置やミトコンドリアの分裂・融合に関わるタンパク質群などが存在し、ミトコンドリアを構成する全タンパク質のおよそ2割(150以上)が含まれている。
タンパク質とリン脂質の重量比は3:1ほどである。
内膜の進化的起源は共生細菌の細胞膜を由来としており、内膜に特徴的なリン脂質カルジオリピンの存在がその証左と考えられている。
一般的に内膜は内側へ向かって陥入し、クリステと呼ばれる構造を形成している。
これによって内膜の表面積の増大、ひいてはATP合成能の増大に寄与している。
外膜と内膜の表面積の比は細胞のATP需要と相関しており、肝臓では5倍ほど、骨格筋ではさらに大きな値である[7]。
クリステの形状は生物によって様々であり、平板状、管状、団扇状、などが知られている。
多細胞動物や陸上植物ではミトコンドリアの長軸に直交する平板状をしており、日本では、教科書などを通じて広く知られている形状である。
しかし、これはむしろ特殊な形状であり、真核生物全体を見渡すと、管状のクリステが一般的である[8]。
さらに、同一個体であっても、組織によってクリステの形状が異なる場合がある。
例えば、ヒトの多くの細胞のミトコンドリアのクリステは平板状だが、副腎皮質や精巣や卵巣でステロイドホルモン類を分泌する細胞が有するミトコンドリアのクリステは、管状や小胞状であったりする[9]。
他にも、ラットでも、このような組織によって、ミトコンドリアのクリステの形状が異なっていることが確認された[10]。
さらには、哺乳類のステロイドホルモン分泌細胞以外でも、平板状だけでなく、管状や小胞状のクリステも有するミトコンドリアが観察される場合もある[11]。
これらのように例外も数多い。
マトリクス
内膜に囲まれた内側がマトリクスであり、TCAサイクル(別名:TCA回路・クレブス回路・クエン酸回路・クエン酸サイクル)やβ酸化など、ミトコンドリアの代謝機能に関わる酵素群が数多く存在している。
ここにはmtDNAが含まれており、ミトコンドリア独自の遺伝情報が保持されている。
その遺伝子発現を担うために、リボソーム、tRNA、転写因子や翻訳因子なども存在している。ミトコンドリア全タンパク質の6割から7割が存在しており、非常にタンパク質濃度の高い区画である。
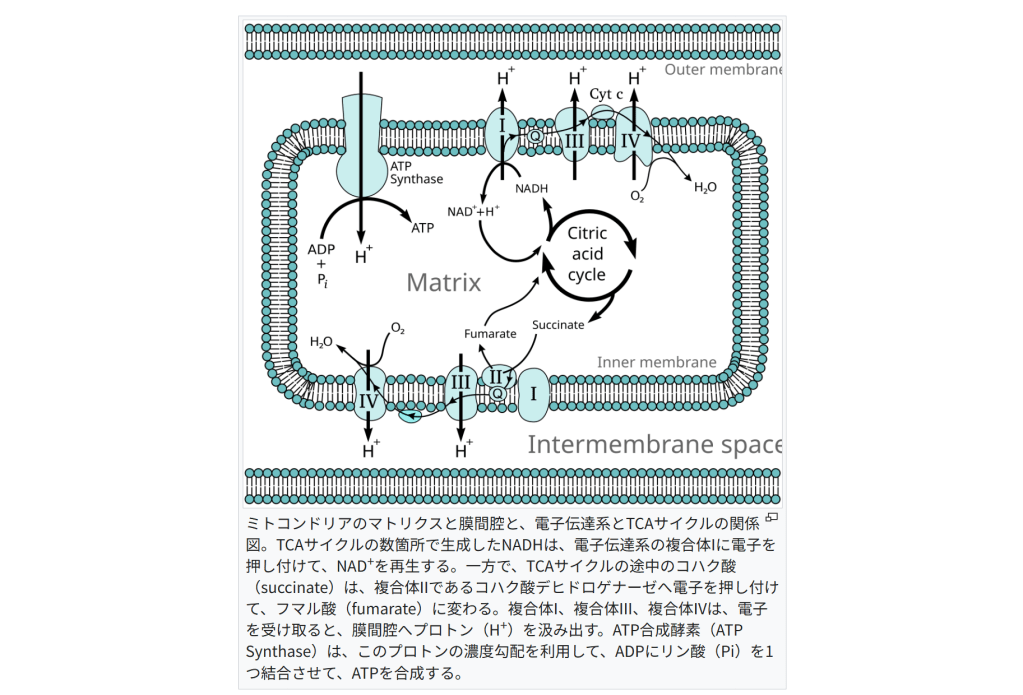
ミトコンドリアのマトリクスと膜間腔と、電子伝達系とTCAサイクルの関係図。
TCAサイクルの数箇所で生成したNADHは、電子伝達系の複合体Iに電子を押し付けて、NAD+を再生する。
一方で、TCAサイクルの途中のコハク酸(succinate)は、複合体IIであるコハク酸デヒドロゲナーゼへ電子を押し付けて、フマル酸(fumarate)に変わる。
複合体I、複合体III、複合体IVは、電子を受け取ると、膜間腔へプロトン(H+)を汲み出す。
ATP合成酵素(ATP Synthase)は、このプロトンの濃度勾配を利用して、ADPにリン酸(Pi)を1つ結合させて、ATPを合成する。
機能
ミトコンドリアの主要な機能は、解糖系やTCAサイクルなどで生成した産物を利用して、電子伝達系に高エネルギーの電子を送り込み、それを酸素に押し付けながら作り出したプロトンの濃度勾配で、ATP合成酵素を駆動して、ADPを酸化的リン酸化によってATPに変換する機能である。[注釈 2]。
もちろん、ミトコンドリアが関与しない解糖系のようなATP産生系も存在するものの、真核生物の細胞の活動に必要なATPの多くは、直接、あるいは間接的にミトコンドリアからATPの形で供給される。
さらに、ミトコンドリアで行われる、TCAサイクル自体でも実質上はATPと等価なGTPも産生されるなどするため、比喩的に「真核細胞のエネルギーを作り出す場」などと説明される場合もある[注釈 3]。
ただし、ATPやGTPの合成以外にもミトコンドリアは多様な機能を有している。
例えば、ステロイドやヘムの合成などを含む様々な代謝、カルシウムや鉄の細胞内濃度の調節、細胞周期やアポトーシスの調節などにも大きく関わっているとされる。
しかし、これらの機能を全てのミトコンドリアが担っている訳ではなく、機能によっては、特定の細胞でのみ動いている。
こうした様々な機能には多数の遺伝子が関わっており、それらに関わる遺伝子の変異が発生した細胞が自然免疫で排除されないと、ミトコンドリアの機能低下を招き、ミトコンドリア病を引き起こす場合がある。
また、ミトコンドリアには他のミトコンドリアの修復作用もあり、近年は細胞から他の細胞へも移動すると考えられ、それに着目した病気の治療法の研究開発も行われている[12]。
ATP産生
ATP産生はミトコンドリアの主たる機能であって、これに関わる多くのタンパク質が内膜やマトリクスに存在している。
細胞質では解糖系が行われ、主にグルコースを代謝して、わずかなATPを合成しながら、ピルビン酸とNADHに分解する。
ここで、もし酸素が充分に存在しない場合には、解糖系の産物は嫌気呼吸により代謝される。
しかしミトコンドリアで酸素を用いて、これらを酸化する好気呼吸を行えば、嫌気呼吸と比べて効率良くATPを得られる。
嫌気性分解では1分子のグルコースから2分子のATPしか得られなかったのに対して、ミトコンドリアによる好気性分解によって、1分子のグルコースから約38分子のATPが合成できる[8][注釈 4]。
また、ミトコンドリアでは、ピルビン酸だけでなく、脂肪酸も利用できる。
ミトコンドリアで脂肪酸はβ酸化が行われる。
ピルビン酸がアセチルCoAに変換されて、TCAサイクルに入るように、β酸化によって、脂肪酸は炭素鎖が2つずつ切り離されてアセチルCoAが生成され、同じようにTCAサイクルに入るからである。
なお、植物のミトコンドリアは、酸素が無くとも、亜硝酸を利用してある程度のATP産生が可能である[13]。
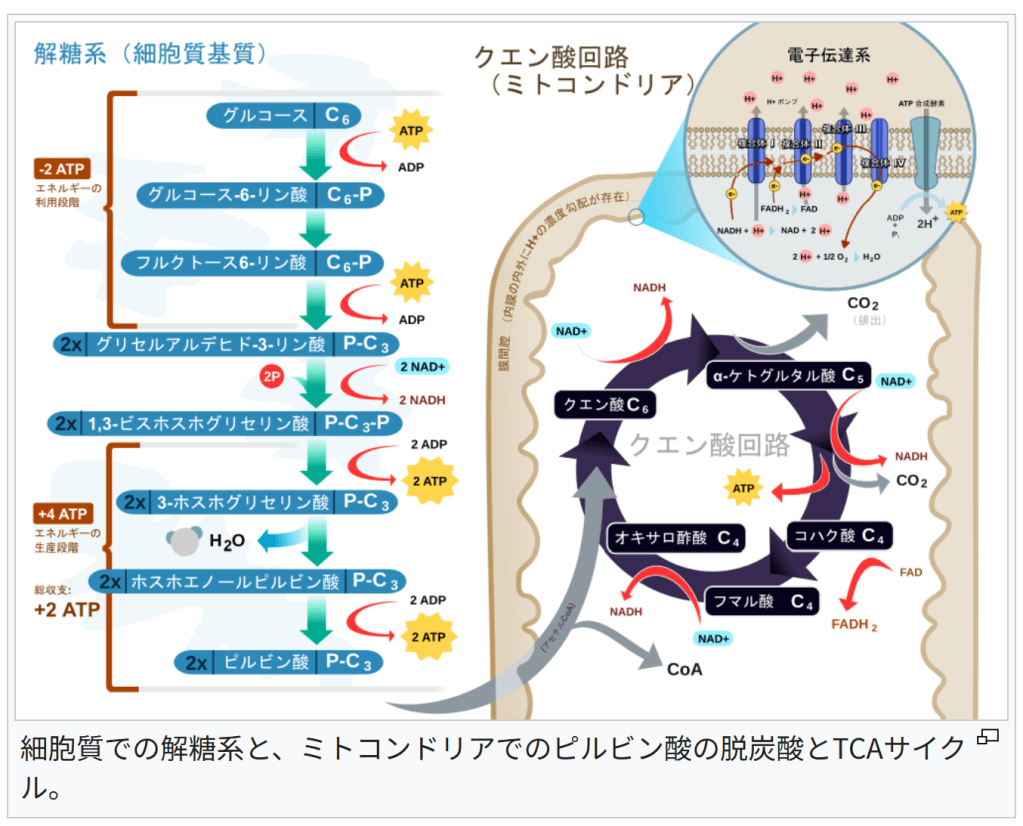
細胞質での解糖系と、ミトコンドリアでのピルビン酸の脱炭酸とTCAサイクル。
細胞質での解糖系
→詳細は「解糖系」を参照地球上の全ての生物で解糖系は、その反応が細胞質基質で起こる。
これは解糖系が細胞内小器官が発生する以前から存在してきた、最も原始的な代謝系であることを反映しているのだろうと考えられている。
真核生物では、解糖系で得られた物質(ピルビン酸とNADH)を、TCAサイクルや電子伝達系の反応を行うミトコンドリアへ輸送し、好気呼吸を行う[14][信頼性要検証]。
ミトコンドリアへの輸送
→詳細は「リンゴ酸-アスパラギン酸シャトル」を参照細胞質の解糖系で生成されたピルビン酸は、ピルビン酸共輸送体(ピルビン酸/H+)により細胞質からミトコンドリアへ輸送される。
同じく細胞質で生成されたNADHはリンゴ酸-アスパラギン酸シャトルによりミトコンドリアへ実質的に輸送される[15]。
ただし、グリセロリン酸シャトルで輸送される場合もあり、この場合にNADHは、ミトコンドリアのTCAサイクルで発生するFADH2相当に、ミトコンドリアでのATPの産生量は目減りする[16]。
なおADPは、ATP/ADPトランスポーターにより細胞質からミトコンドリアへ輸送される[17]。
また、H2O、O2、CO2、NH3は、そのままミトコンドリア内膜を通過できる。
ピルビン酸の脱炭酸
アセチルCoAは、好気性細胞呼吸の第2段階目である、ピルビン酸がピルビン酸デヒドロゲナーゼ複合体により脱炭酸して生成する。この酵素反応はミトコンドリアのマトリクスで起こる。ここで生成したアセチルCoAは、TCAサイクルに投入される[18][信頼性要検証]。
TCAサイクル
→詳細は「TCAサイクル」を参照解糖系で生じたピルビン酸は内膜を能動輸送によって透過し、マトリクスで酸化され補酵素Aと結合し、二酸化炭素、アセチルCoA、NADHを生じる。
アセチルCoAは、TCAサイクルへ入る基質である。
TCAサイクルの反応に関わる酵素群は、ほとんどがミトコンドリアのマトリクスに存在している。
しかし、コハク酸デヒドロゲナーゼだけは例外で、内膜の内側に付着しており、これが電子伝達系の複合体IIに当たる。
TCAサイクルで、コハク酸からフマル酸に変換する際の酸化還元反応では、電子伝達系の複合体Iを動かすほどのエネルギーが無く、複合体IIが動かされる。
その後は、いずれも電子伝達系の複合体IIIへとエネルギーが伝達され、ATP産生に寄与する。
なお、TCAサイクルはアセチルCoAを酸化して二酸化炭素を生じ、その過程で3分子のNADHと1分子FADH2、1分子のGTPを生成する。
なお、二酸化炭素はミトコンドリア外へ拡散して排出される。
TCAサイクルでは、サイクルの1回転ごとに、全ての中間体(例えば、クエン酸、イソクエン酸、α-ケトグルタル酸、コハク酸、フマル酸、リンゴ酸およびオキサロ酢酸)が再生される。
したがって、ミトコンドリアにこれらの中間体のいずれかを追加して加えると、追加された量がTCAサイクル内に保持され、中間体の1つが他方に変換されて順次増加する。
よって、それらの中間体のいずれか1つをTCAサイクルに加えれば、補充反応(アナプレロティック反応)効果を示す。
逆に、中間体のいずれかの除去すれば、消費反応(カタプレロティック反応)効果を示す。
これらの補充反応及び消費反応は、TCAサイクルの回転で、アセチルCoAと結合してクエン酸を形成するために利用可能な、オキサロ酢酸の量を増加または減少させる。
この回転量が、ミトコンドリアによるATP製造量と、細胞へのATPの提供量の増減を左右する[19]。
要するに、ミトコンドリア内に存在するTCAサイクルの各種中間体の量が、TCAサイクルの反応速度を調節し、ATPの合成量も調節することを意味する。
電子伝達系
→詳細は「電子伝達系」および「酸化的リン酸化」を参照NADHやFADH2が有する還元力は、内膜にある電子伝達系で数段階を経て、最終的に酸素に渡される。要するに、電子を、電気陰性度の高い酸素に押し付ける形である。
なおNADHは、マトリクスでのTCAサイクルやβ酸化だけでなく、細胞質の解糖系でも生ずる。
細胞質で生じたNADHの還元力は、マロン酸-アスパラギン酸対向輸送系や、リン酸グリセロールシャトル系を通じて電子伝達系に供給される。
内膜の電子伝達系には、NADH脱水素酵素、シトクロームc還元酵素、シトクロームc酸化酵素が存在しており、プロトン(H+)を膜間腔へ汲み出す。
この過程は非常に効率的だが、不充分な反応により活性酸素種を生じ得る(活性酸素#活性酸素と人体の関係参照のこと)。
これがいわゆる「酸化ストレス」の形態の1つであり、ミトコンドリアの機能低下や老化に関与していると考えられている[20]。
グルコーストランスポーターであるGLUT1を介して、デヒドロアスコルビン酸がミトコンドリアに輸送され、その後アスコルビン酸に還元され、活性酸素によるフリーラジカルの大部分が生成される場所であるミトコンドリアに蓄積される。
アスコルビン酸は、ミトコンドリアの脂質膜とmtDNAを、活性酸素による酸化から保護する[21]。
電子伝達系で、複合体Iと複合体IIIと複合体IVは、電子が伝達された際に、ミトコンドリアのマトリクスから膜間腔へとプロトンを汲み出す。
このようにしてプロトンが膜間腔へ汲み出された結果、ミトコンドリアの内膜の隔てて、プロトン濃度の差(電気化学的勾配)が生じる。
汲み出されたプロトンは、ATP合成酵素を通じてマトリクスへ戻ることができ、この際に、電気化学的勾配のポテンシャルを使って、ADPと無機リン酸(Pi)を、ATPへと変換する。
生成されたATPは、ATP/ADPトランスポーターによって、ミトコンドリアから細胞質へ輸送され、細胞の活動エネルギー源として利用される[17]。この原理を化学浸透説と呼び、これをピーター・ミッチェルが最初に唱えた功績によって、1978年にノーベル化学賞を受賞した[22][23]。
また、ATP合成酵素の反応機構を明らかにしたポール・ボイヤーとジョン・E・ウォーカーには、1997年にノーベル化学賞が授与された[24]。
脂肪酸の輸送とβ酸化
ミトコンドリアにおける脂肪酸の輸送については、β酸化#脂肪酸の動員及びβ酸化#脂肪酸の活性化とミトコンドリア内への輸送を参照のこと。ミトコンドリアにおけるβ酸化については、β酸化#β酸化反応および酵素群を参照のこと。
なお、このミトコンドリアのマトリクスで行われる脂肪酸のβ酸化によって、1分子のアセチルCoAを生成する反応の際に、1分子のATPを消費するものの、FADH2とNADHと1分子ずつ生成する。
このFADH2とNADHは、電子伝達系に使用され、より多くのATPを産生できる。
さらに、ミトコンドリアのマトリクスで生成されたアセチルCoAは、同じくマトリクスで行われているTCAサイクルに投入され、さらに、GTPやATPを産生できる。
アセトアルデヒドの酸化
ミトコンドリアにはアルデヒドデヒドロゲナーゼも発現している[25]。
飲酒などによってエタノールを体内に摂取すると、肝臓などで発現しているアルコールデヒドロゲナーゼなどの作用によって代謝され、アセトアルデヒドが生成する。
このアセトアルデヒドを、ミトコンドリアはアルデヒドデヒドロゲナーゼで代謝して、酢酸に変換できる。
このミトコンドリアでのアルデヒドデヒドロゲナーゼの活性が遺伝的に低いヒトが、東洋人などの一部に見られ、そのようなヒトはアセトアルデヒドの毒性が強く出やすい[25]。
なお、酢酸とは、炭素鎖2つの脂肪酸である。筋肉とミトコンドリア
速筋線維はミトコンドリアが少なく、グリコーゲンが比較的多いので白く見える。
解糖系でATPを産生し、その結果として蓄積したピルビン酸は、乳酸デヒドロゲナーゼで乳酸へと変換されやすい。
このような嫌気的な糖分解によるATP産生であれば、わざわざ外部から酸素を取り込む必要もなく、速くATPを作り出せる。
このこともあり、乳酸性閾値よりも高い運動強度では、速筋線維が多く使われるようになる。
しかしながら、この方法では長時間の運動は続けられないという欠点がある。
これに対して、遅筋線維や心筋は、ミオグロビンが多いので赤く見え、酸素を利用しやすい環境を備えている。
赤色の筋肉では、乳酸を作るよりは、解糖系の産物であるピルビン酸をミトコンドリアのTCAサイクルへ、解糖系で生成したNADHもミトコンドリアに渡され、ATPを合成して、運動のために使っている。
この方式であれば、乳酸などが蓄積しないので、運動強度が低い場合は遅筋線維が主として働いている。
なお、速筋線維で発生した乳酸は、血液を介して肝臓に運ばれ、コリ回路でATPを消費してグルコースの再生に使われることは、よく知られている。
これ以外に、乳酸デヒドロゲナーゼは、乳酸をピルビン酸に戻す逆反応も触媒できる。
遅筋線維や心筋では、外部から取り込んだ乳酸を、ピルビン酸に戻して、ミトコンドリアのTCAサイクルに投入することも行っている。
いずれにしても、主に速筋線維で蓄積しやすい乳酸の代謝には、細胞膜を通過して他の細胞へと乳酸が輸送される必要がある。
この乳酸の輸送は、乳酸だけでなくピルビン酸などの輸送にも関わるため、モノカルボン酸の輸送担体(英語: Monocarboxylate Transporter (MCT))と呼ばれている[26]。
熱産生
ある条件下では、膜間腔のプロトンはATP合成に関与せずに、促進拡散によってマトリクスに戻る場合がある。
これは「プロトンのリーク」とか「ミトコンドリアの脱共役」と呼ばれ、これによって蓄積されていた電気化学ポテンシャルは熱として解放される。
サーモジェニン(英語版)などの一群のプロトンチャネル(脱共役タンパク質)が媒介しており、筋肉の震えを伴わない熱産生に関わっている[27]。
サーモジェニンは、若齢や冬眠中の哺乳類に見られる褐色脂肪組織のミトコンドリアに多く存在している。
アポトーシス
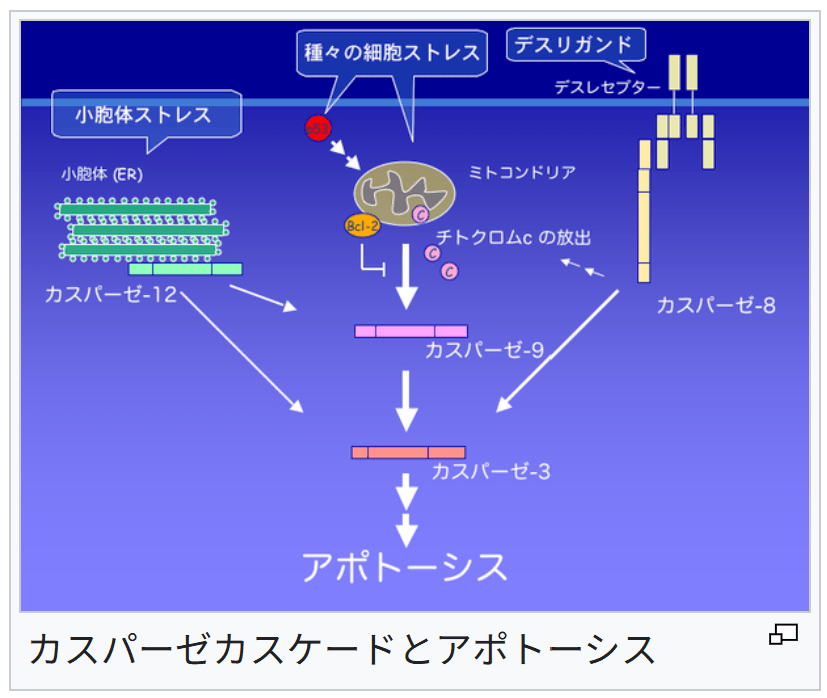
カスパーゼカスケードとアポトーシス
→詳細は「アポトーシス」および「カスパーゼ」を参照細胞に発生したDNA損傷などのストレスは、アポトーシス誘導分子p53やアポトーシスを調節するBcl-2ファミリータンパク質を介して、ミトコンドリアの膜電位を変化させ、外膜の電位依存性陰イオンチャネルが閉鎖される[28]。
なお、ミトコンドリアの外膜の電位依存性陰イオンチャネルが閉鎖されると、ミトコンドリアの機能は低下する。
さらに、ミトコンドリアの膜電位の変化は、ミトコンドリアからのシトクロムcの漏出も発生させ、アポトーシスへとつながる[29]。
シトクロムcは、細胞質に存在するApaf-1やカスパーゼ-9と結合して、アポトソーム(英語: apoptosome)と呼ばれる集合体を形成する。
これによって活性化されたカスパーゼ-9が、下流のエフェクターを活性化する。
この後は、DNAが切断されて、細胞は自殺する。
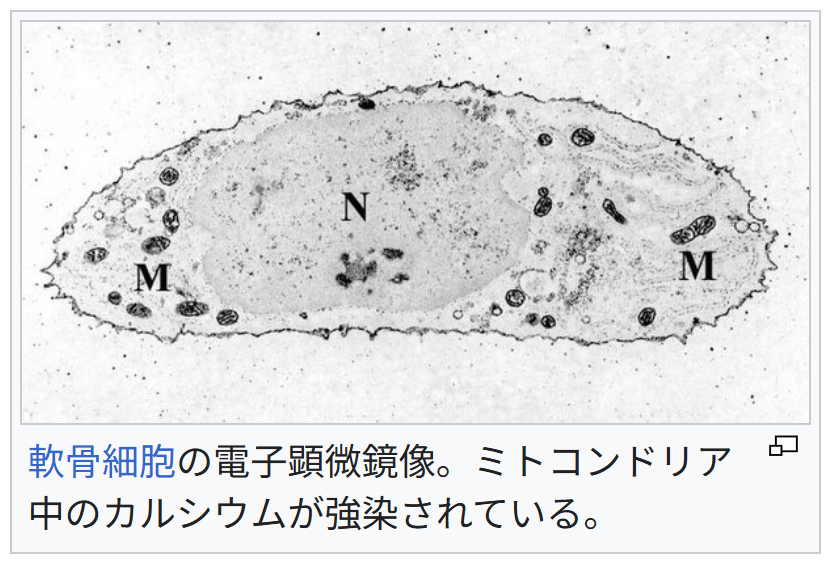
カルシウム貯蔵
軟骨細胞の電子顕微鏡像。ミトコンドリア中のカルシウムが強染されている。
細胞内のカルシウム濃度は様々な機構によって厳密に制御されており、細胞中の情報伝達に重要な役割を果たしている。
細胞内のカルシウム濃度の上昇により、セカンドメッセンジャー系が起動されたり、筋肉の収縮が起きたりと、様々な反応が起きる。
細胞内におけるカルシウムの貯蔵場所としては小胞体が最も顕著だが、カルシウムの貯蔵に関して、小胞体とミトコンドリアは協調している[30]。
というのも、ミトコンドリアは一過的なカルシウム貯蔵能を有し、細胞におけるカルシウム濃度の恒常性に貢献しているのである[31]。
ミトコンドリアは迅速にカルシウムを取り込むことが可能で[32]、カルシウムは内膜のカルシウム輸送体により、マトリクスへと取り込まれる[33]。これの動作は、ミトコンドリアの膜電位に依存している[31]。
こうして取り込んだカルシウムを、ミトコンドリアが後々放出することで、カルシウム濃度の緩衝作用を果たしている[32][34][35]。
なお、カルシウムの放出は、ナトリウム・カルシウム対向輸送、もしくは、カルシウム依存性カルシウム放出系によって行われる[33]。
ミトコンドリアゲノム
→詳細は「ミトコンドリアDNA」を参照ミトコンドリア中には、細胞核とは別に、独自のDNAが存在しており、これをミトコンドリアDNA(mtDNA)と呼ぶ。
mtDNAは、細胞核とは異なる独自の遺伝情報を持っている。
DNA分子の大きさや形状、codeされている遺伝子の数や種類などは、生物種によって大きく異なる。
ただ、通常はGC含量が低く(20-40%)、基本的なmtDNAは、塩基対が数十kb程度のDNAである。
mtDNAには、電子伝達系に関わるタンパク質、リボソームRNAやtRNAなど、数十種類の遺伝子がcodeされている。
ヒトを含む脊椎動物のmtDNAは、真核生物の中ではかなり特殊な性質を多く持っており、研究はよく進んでいるものの、安易な一般化は慎まなければならない。
なお、mtDNAと、それに基づいて合成される産物の一部は、ミトコンドリアだけではなく、細胞表面にも所在し、mtDNAに突然変異が発生している場合には、自然免疫系が特異的に細胞ごと破壊して排除する[36]。
mtDNAに突然変異が発生した場合には、ミトコンドリア病を発症する可能性もある。
mtDNAの塩基対数と形状
最も小さなmtDNAを持つ生物はアピコンプレックス門の原虫で、大きさわずか6 kbの線状ゲノムである。
電子伝達系に関わる3つのタンパク質遺伝子と、断片化されたリボソームRNA遺伝子群のみが存在している。
逆に最も大きなmtDNAは、マスクメロンの持つ2400 kbという巨大なゲノムである。
ただし遺伝子数は比較的多いものの、それでも100弱に過ぎず、大量の反復配列やグループ2イントロンなどの非遺伝子領域が大部分を占める。
ヒトを含む多細胞動物のmtDNAはいずれも比較的似通っており、長さ16 kb前後の単一の環状DNAで構成されている。
遺伝子は37あり、その内訳は、呼吸鎖複合体とATP合成酵素のサブユニットが13、tRNAが22、rRNAが2である。
遺伝子地図などでは、mtDNAが環状に表現される事例が多い。
しかし物理的に環状のmtDNAを持つ生物はごく一部に限られ、多くの生物では環状の基本構造からトイレットペーパーを引き出すかのように連続的に複製されており、その結果mtDNAの大部分は、基本単位が何度も繰り返す線状反復構造を有している。
また少数派ではあるものの、常に線状のmtDNAを持つ生物も存在している。
遺伝子
ミトコンドリアゲノムはαプロテオバクテリアから受け継がれており、その遺伝子発現は細菌と共通した特徴を持っており、真核生物の細胞核のDNAとは異なる。
例えば、複数の遺伝子がまとめて転写され、それが遺伝子ごとに切断されポリアデニル化されて成熟mRNAとなる点や、翻訳の開始にフォルミル化メチオニンが利用される点、細胞核に存在するようなスプライソソーム型のイントロンが存在しない点、などが挙げられる。
さらに、ミトコンドリアの遺伝暗号表は、細胞核や一般の原核生物で利用されている普遍暗号表と比べて、若干の差が見られる。
顕著な例として、細胞核では終止コドンであるはずのUGAが、ミトコンドリアではトリプトファンをコードしている場合が多いことが挙げられるものの、例外も多く、生物種によって少しずつ異なる暗号表を用いているのが実態である[37]。
またミトコンドリアでは、しばしばRNA編集が行われる。
例えば高等植物のミトコンドリアでは、DNA配列上のCGGがmRNAの場合は、UGGと編集されてトリプトファンをコードするという例が知られている[38]。
ただ、重要な点として、ミトコンドリアの機能に関わる全ての遺伝子が、mtDNAに存在しているわけではないが挙げられる。
ミトコンドリアが持つmtDNA上にcodeされているミトコンドリアゲノムは、細菌のゲノムと比べると、遺伝子数が極端に減少している。
一方で、ミトコンドリアが必要とする大多数の遺伝子は、細胞核の側にcodeされており、細胞質の側で転写された情報に基づいて生合成された遺伝子産物が、ミトコンドリアへと輸送される。
これは進化の過程で、遺伝子が細胞核へ移動したからだと考えられている。
こうした現象は、比較的よく起きた出来事だと考えられ、マイトソームなどのように全てのDNAを完全に失ったようなミトコンドリアも存在している。
一方で、原生生物のレクリノモナス(Reclinomonas americana)は、他の生物では細胞核から輸送されているようなタンパク質の遺伝子が、mtDNA上に存在しており、比較的原始的なミトコンドリアゲノムを未だに保持していると考えられている。
ミトコンドリアには、呼吸機能に関与する疎水性のタンパク質が存在し、疎水性であるために輸送が難しく、これらをミトコンドリアの内部で作らざるを得ないために、ミトコンドリアに遺伝子が残っている理由の1つと考えられている[39]。
異数性
1つのミトコンドリアには、2-10コピーのDNA分子が存在する[40]。
その全てが完全に同じ情報を持つわけではなく、複数の異質のDNA分子を含んでいると確認されている[8]。
起源
ミトコンドリアは、リケッチアに近い好気性細菌のαプロテオバクテリアが、約20億年ほど前に真核細胞に入り込んだ結果として獲得されたと考えられている[8][41]。
リン・マーギュリスの細胞内共生説では単に好気性バクテリアが起源とされていたが、その後すぐの1970年代にはすでにミトコンドリアの起源が現在でいうαプロテオバクテリアだという意見が出た。
脱窒細菌のParacoccus denitrificansや、暗所好気条件で培養した紅色光合成細菌のRhodobacter sphaeroidesは、呼吸鎖の構成や阻害剤への応答がミトコンドリアと類似しており、特に、シトクロムcがミトコンドリアと互換性を持つ点が注目された[42]。
二重の膜があるが、一つが元のバクテリアの本来の膜、もう一つがバクテリアが細胞内に入り込む際にそれを包み込んだ細胞自体の膜だったと考えられている。
細胞核DNAにコードされているシトクロムcだけでなく、mtDNAにコードされているリボソームRNAの配列を使った系統解析でもαプロテオバクテリア起源であると示され、1980年代にはミトコンドリアのαプロテオバクテリア起源は受け入れられるようになった[43]。
ただし、初期の解析では高等植物ミトコンドリアのリボソームRNAの配列が、他のミトコンドリアの配列と比べて進化的距離が非常に小さかったため、ミトコンドリアの起源は単独ではなく、高等植物のミトコンドリアは新たに獲得された物だという意見もあった[43]。しかし、こうした意見は現在では否定され、真核生物のミトコンドリアの起源は単一であるとされている[44]。
もっとも、αプロテオバクテリアは非常に多様な細菌を含む分類群であり、その中でどのような細菌がミトコンドリアの起源なのかについては、長く議論が続いている。
初期には前述の通り脱窒細菌や光合成細菌が起源だと考えられていたが、シャペロニン(英語版)Hsp60(GroEL)を用いた系統解析によりリケッチアが最も近縁であると示されてからは、これが有力説となった。
リケッチアは、細胞内寄生生物である点、TCAサイクルを持ち好気呼吸ができるのに対して解糖系を持たない点、細胞膜にADP/ATP輸送体を持っている点、ゲノムが小さくAT含量が高い点など、ミトコンドリアと共通した特徴が複数見られる[44]。
1998年に発疹チフスを引き起こすリケッチアの1種であるRickettsia prowazekiiのゲノムが解読され、祖先的とされるReclinomonas americanaのミトコンドリアゲノムと共通している遺伝子や、配置順が保存された遺伝子群などが見出され解析された。
その多くはミトコンドリアがリケッチアに近縁であるという仮説を支持する結果であったが、ADP/ATP輸送体については、予想に反して起源を異にしていると示された[45]。
20世紀末から21世紀初頭にかけて、世界中の海洋には自由生活性で浮遊性の細菌ペラジバクター(暫定的にCandidatus Pelagibacter ubiqueと命名されている)が存在していることが明らかとなった[46]。
ペラジバクターはリケッチア目の中で、最も祖先的な位置から派生したと考えられる生物であり、ミトコンドリアの起源を、ペラジバクターとその他一般的なリケッチアとの間に求められる[47]。
なお、アメーバに似た原生生物であるペロミクサや微胞子虫など、原生生物の中はミトコンドリアを持っていないものもいる。
これを、ミトコンドリアが共生する以前の真核生物の生き残りと見る説があった(→アーケゾア仮説)が、後に否定された。
生物の系統との関係
ミトコンドリアの特徴は、動物、植物、菌類にほぼ共通であるが、それ以外の原生動物では、若干異なった形状の物がある。
特にクリステの形については、明らかに異なった形状のミトコンドリアが見られる。
ヒトなどの一般のミトコンドリアでは、内膜がひだのように折れ曲がり、クリステは平坦な板のような形をしている。
しかし、粘菌類の場合、クリステは内膜から内部へと放射状に入り込む管の形で、管の表面にATP合成酵素の手段が並んでいる。
また、内部の中央にDNAを含んだ塊があって、ミトコンドリア核と呼ばれる。
このような、管状のクリステを持つミトコンドリアは、繊毛虫やアピコンプレックス類、アメーバ類、クロララクニオン藻類などの原生生物にも見られる。
また、ミドリムシ類とトリパノソーマでは、クリステは団扇型である。
これらのミトコンドリアは、細長くて枝分かれをして、細胞内に広がっている。
トリパノソーマでは、鞭毛の基部にキネトプラストと呼ばれる袋状の構造が知られており、その中の顆粒にはDNAが含まれているが、これはミトコンドリアの一部である。
がんとミトコンドリア
1955年にオットー・ワールブルクは、体細胞が長期間低酸素状態に晒されると呼吸障害を引き起こし、通常の酸素濃度の環境下に戻しても、大半の細胞が変性や壊死を起こし、ごく一部の酸素呼吸に代わるエネルギー生成経路を昂進させて生存する細胞が、ガン細胞になるとの説を発表した。
この説では、酸素呼吸よりも、むしろ解糖系によるエネルギー産生に依存する細胞は、下等動物や胎生期の未熟な細胞が一般的であり、体細胞がATP産生を酸素呼吸によらず解糖系に依存した結果、細胞が退化してガン細胞が発生するとした[48]。
ガン細胞の発生とmtDNAの突然変異の関与は、古くから指摘されてきた。
その理由は特定の発ガン性化学物質が、DNAよりもmtDNAに結合しやすいことと、ガン組織のmtDNAは正常組織よりも高い割合で突然変異の蓄積が観察されたことによる。
しかしながら、母性遺伝するガンの存在が確認されていない点や、DNAの影響を排除しmtDNA単独でのガンへの影響を検証する手法が確立されていない点などが、この仮説の証明の障害であった。
ただ、2008年筑波大学の林純一らが、ガンの転移能獲得という、ガン細胞の悪性化に、mtDNAの変異が関与していることを指摘した[49]。
マウス肺がん細胞の細胞質移植による細胞雑種の比較により、mtDNAの特殊な病原性突然変異によってガン細胞の転移能獲得の原因になることを発見し、ヒトのガン細胞株でも、mtDNAの突然変異がガン細胞の転移能を誘導し得ることを明らかにし、少なくとも、mtDNAがATP合成以外の生命現象にも関与することを明らかにした。
また、林らによるとmtDNAの突然変異には、活性酸素種(ROS)の介在が重要であり[49]、ROSを除去すれば転移能の抑制が可能ではないかと推察した。
ただし、ガン細胞の転移能の獲得メカニズムは複雑であり、様々な要因が考えられるので、これはその要因の1つに過ぎない。
また、京都大学の井垣達吏らは、① Ras遺伝子の活性化とミトコンドリアの機能障害を起こした細胞は、細胞老化を起こして細胞老化関連分泌因子(SASP因子)を放出し、これにより周辺組織のガン化を促進すること、
また、② 細胞分裂停止とJNK遺伝子の活性化が互いに増幅し合うことで、細胞内のJNK活性が顕著に増大し、これによりSASP因子の産生が誘導されることを示した[50][51][52]。
「ミトコンドリア・イブ」
→詳細は「ミトコンドリア・イブ」を参照ミトコンドリアのDNAは、同種交配の場合卵子に入った精子のミトコンドリアが選択的に排除されるため、母親のmtDNAを引き継ぐことを根拠に、現生人類の起源の地が探られた。
すなわち、世界中に分布するヒトからmtDNAを調べて、現在の分布地図から現生人類の起源とその移動について推察する作業を実施した。
この結果、大昔のアフリカのある女性が、今の人類の全てのミトコンドリアについての「母親」であるとの仮説が発表された。
この女性はキリスト教徒の宗教的説話になぞらえて「ミトコンドリア・イブ」と呼ばれている。
しかしながら、この仮説は、その他の遺伝情報について、この女性に全てが由来するという意味ではない。
無論、全人類の起源が1人の女性にあると言っているわけでもない。
しかも、実験的に異種交配させた受精卵では、精子由来のミトコンドリアを排除するプロセスが失敗する場合があることが知られている。
フィクション
小説
1995年に第2回日本ホラー小説大賞を受賞した瀬名秀明の『パラサイト・イヴ』は、ミトコンドリアの共生起源説、および、人類の進化におけるミトコンドリア・イブ説に基づき、現在のミトコンドリアは細胞の支配下にあるが、もしもそれが反乱を起こしたならば、という仮定の物語で話題を呼び、映画やゲーム化も行なわれた。
脚注
[脚注の使い方]注釈
^ ミトコンドリアのマトリクス(mitochondrial matrix)は、マトリックスと片仮名転記される場合もある。さらに、ミトコンドリア基質(mitochondrioplasma)とも呼ばれる。ただ、本稿では「マトリクス」の表記で統一する。
^ したがって、これが阻害されると、真核生物の細胞は深刻なATP不足に陥り得る。例えば、シアン化水素や硫化水素などが毒である理由は、ミトコンドリアの電子伝達系の複合体IVを阻害するためである。他にも、電子伝達系の複合体Iを阻害するアモバルビタールなど、電子伝達系の複合体IIを競合的に阻害するマロン酸など、電子伝達系の複合体IIIを阻害するジメルカプロールなど、ATP合成酵素を阻害するオリゴマイシン(英語版)など、ここに関わる物質は多数存在する。なお、これらとは別に、2,4-ジニトロフェノールのような、電子伝達系とATP合成酵素の作用を切り離してしまう脱共役剤と呼ばれる毒物も存在する。ただし、体温を上昇させるために、敢えて生体が制御した脱共役を行うためのサーモジェニン(英語版)と呼ばれるタンパク質も存在する。つまり、生理的な条件下でも、わざと脱共役が行われる場合もある事が知られている。
^ しかしながら、これは比喩であって、ミトコンドリアがエネルギーを作り出しているわけではない。あくまで、外来の高エネルギーの物質を、細胞が活動する際に使い易い、ATPやGTPなどの形に変換しているだけである。この際に、ロスも出るため、実質的なエネルギーは、減少している。
^ ミトコンドリアのシャトル系などの関係で、多少の変動が出る。なお、この1分子のグルコースから、約38分子のATPという比率は、代謝系に阻害が行われておらず、かつ、サーモジェニン(英語版)などが動いていない場合の話である。出典
^ ニック・レーン(著)斉藤隆央(訳)『ミトコンドリアが進化を決めた』 p.1、P.16、みすず書房、2007年、ISBN 978-4-622-07340-6
^ “mitochondria”. Online Etymology Dictionary. 2018年11月27日閲覧。
^ a b c 藤田 尚男・藤田 恒夫 『標準組織学 総論(第3版)』 p.50 医学書院 1988年2月1日発行 ISBN 4-260-10047-5
^ Hayashi T.; Rizzuto R.; Hajnoczky G.; Su TP. (February 2009). “MAM: more than just a housekeeper”. Trends Cell Biol. 19 (2): 81-88. doi:10.1016/j.tcb.2008.12.002. PMC 2750097. PMID 19144519.
^ Herrmann JM.; Neupert W. (April 2000). “Protein transport into mitochondria”. Curr Opin Microbiol 3 (2): 210-214. doi:10.1016/S1369-5274(00)00077-1. PMID 10744987.
^ Chipuk JE.; Bouchier-Hayes L.; Green DR. (2006). “Mitochondrial outer membrane permeabilization during apoptosis: the innocent bystander scenario”. Cell Death and Differentiation. 13 (8): 1396-1402. doi:10.1038/sj.cdd.4401963. PMID 16710362.
^ Mannella CA (2006). “Structure and dynamics of the mitochondrial inner membrane cristae”. Biochimica et biophysica acta 1763 (5-6): 542-548. doi:10.1016/j.bbamcr.2006.04.006. PMID 16730811.
^ a b c d 黒岩常祥(著)『ミトコンドリアはどこからきたか』 日本放送出版 2000年6月30日第1刷発行 ISBN 4140018879
^ 藤田 尚男・藤田 恒夫 『標準組織学 総論(第3版)』 p.50、p.51 医学書院 1988年2月1日発行 ISBN 4-260-10047-5
^ 藤田 尚男・藤田 恒夫 『標準組織学 総論(第3版)』 p.52 医学書院 1988年2月1日発行 ISBN 4-260-10047-5
^ 藤田 尚男・藤田 恒夫 『標準組織学 総論(第3版)』 p.51 医学書院 1988年2月1日発行 ISBN 4-260-10047-5
^ “NHKスペシャル 人体Ⅲ 第2集 細胞40兆 限りあるから命は輝く”. NHK. 2025年5月25日閲覧。
^ Stoimenova M.; Igamberdiev AU.; Gupta KJ.; Hill RD. (July 2007). “Nitrite-driven anaerobic ATP synthesis in barley and rice root mitochondria”. Planta 226 (2): 465-474. doi:10.1007/s00425-007-0496-0. PMID 17333252.
^ TCA回路 講義資料のページ
^ Monty Krieger; Matthew P Scott; Matsudaira, Paul T.; Lodish, Harvey F.; Darnell, James E.; Lawrence Zipursky; Kaiser, Chris; Arnold Berk. Molecular Cell Biology, Fifth Edition. San Francisco: W. H. Freeman. ISBN 0-7167-4366-3
^ Robert K. Murray・Daryl K. Granner・Victor W. Rodwell(編集)、上代 淑人(監訳)『Illustrated ハーパー・生化学(原書27版)』 p.123 丸善 2007年1月30日発行 ISBN 978-4-621-07801-3
^ a b 真島 英司、寺田 弘、「ATPはいかにして膜を透過するか:ループの協調的スウィングによるミトコンドリアADP/ATPキャリアーの機能発現」、『生物物理』Vol. 38 (1998) No. 6、P 245-249
^ 第5回 5.エネルギーの生産-サイトゾールとミトコンドリア 更新日:2002/04/08 教養部生物学の資料[1]
^ Stryer, Lubert (1995). “Citric acid cycle.”. In: Biochemistry. (4 th ed.). New York: W.H. Freeman and Company. pp. 509-527, 569-579, 614-616, 638-641, 732-735, 739-748, 770-773. ISBN 0-7167-2009-4
^ Huang, K.; K. G. Manton (2004). “The role of oxidative damage in mitochondria during aging: A review”. Frontiers in Bioscience 9: 1100-1117. doi:10.2741/1298. PMID 14977532.
^ KC S.; Carcamo JM.; Golde DW. (2005). “Vitamin C enters mitochondria via facilitative glucose transporter 1 (Glut1) and confers mitochondrial protection against oxidative injury”. FASEB J 19 (12): 1657-1667. doi:10.1096/fj.05-4107com. PMID 16195374.
^ Mitchell P.; Moyle J. (1967-01-14). “Chemiosmotic hypothesis of oxidative phosphorylation”. Nature 213 (5072): 137-139. doi:10.1038/213137a0. PMID 4291593.
^ Mitchell P. (1967-06-24). “Proton current flow in mitochondrial systems”. Nature 25 (5095): 1327-1328. doi:10.1038/2141327a0. PMID 6056845.
^ “Chemistry 1997”. Nobel Foundation (1997年). 2007年12月16日閲覧。
^ a b Robert K. Murray・Daryl K. Granner・Victor W. Rodwell(編集)、上代 淑人(監訳)『Illustrated ハーパー・生化学(原書27版)』 p.247 丸善 2007年1月30日発行 ISBN 978-4-621-07801-3
^ 新たな乳酸の見方、八田 秀雄、学術の動向、Vol. 11 (2006) No. 10
^ Mozo J.; Emre Y.; Bouillaud F.; Ricquier D.; Criscuolo F. (November 2005). “Thermoregulation: What Role for UCPs in Mammals and Birds?”. Bioscience Reports. 25 (3-4): 227-249. doi:10.1007/s10540-005-2887-4. PMID 16283555.
^ Lemasters JJ.; Holmuhamedov E. (2006). “Voltage-dependent anion channel (VDAC) as mitochondrial governator–thinking outside the box.”. Biochim. Biophys. Acta 1762 (2): 181-190. doi:10.1016/j.bbadis.2005.10.006. PMID 16307870.
^ 太田 成男; 石橋 佳朋 (1999). “アポトーシスの分子機構”. 脳と発達 31 (2). doi:10.11251/ojjscn1969.31.122.
^ Pizzo P.; Pozzan T. (October 2007). “Mitochondria-endoplasmic reticulum choreography: structure and signaling dynamics”. Trends Cell Bio. 17 (10): 511-517. doi:10.1016/j.tcb.2007.07.011. PMID 17851078.
^ a b Editor-in-chief, George J. Siegel; editors, Bernard W. Agranoff… [et al.]; illustrations by Lorie M. Gavulic (1999). Siegel GJ, Agranoff BW, Fisher SK, Albers RW, Uhler MD.. ed. Basic Neurochemistry (6 ed.). Lippincott Williams & Wilkins. ISBN 0-397-51820-X
^ a b Rossier MF. (2006). “T channels and steroid biosynthesis: in search of a link with mitochondria”. Cell Calcium. 40 (2): 155-164. doi:10.1016/j.ceca.2006.04.020. PMID 16759697.
^ a b Miller RJ. (1998). “Mitochondria – the kraken wakes!”. Trends in Neurosci. 21 (3): 95-97. doi:10.1016/S0166-2236(97)01206-X.
^ Brighton, Carl T.; Robert M. Hunt (1974). “Mitochondrial calcium and its role in calcification.”. Clinical Orthopaedics and Related Research 100: 406-416.
^ Brighton, Carl T.; Robert M. Hunt (1978). “The role of mitochondria in growth plate calcification as demonstrated in a rachitic model.”. Journal of Bone and Joint Surgery 60-A: 630-639.
^ 林純一 (2011-10-12). “ミトコンドリアDNAに突然変異をもつ細胞は自然免疫により排除されることを発見”. Journal of Experimental Medicine (電子版) 2011.Oct.12.
^ Jukes TH.; Osawa S. (1990-12-01). “The genetic code in mitochondria and chloroplasts”. Experientia. 46 (11-12): 1117-1126. doi:10.1007/BF01936921. PMID 2253709.
^ Hiesel R.; Wissinger B.; Schuster W.; Brennicke A. (2006). “RNA editing in plant mitochondria”. Science. 246 (4937): 1632-1634. doi:10.1126/science.2480644. PMID 2480644.
^ 池田清彦、『不思議な生き物-生命38億年の歴史と謎』 p.203、2013年4月25日、角川学芸出版、ISBN 978-4-04-653275-6
^ Wiesner RJ.; Ruegg JC.; Morano I. (1992). “Counting target molecules by exponential polymerase chain reaction, copy number of mitochondrial DNA in rat tissues”. Biochim Biophys Acta 183 (2): 553-559. PMID 1550563.
^ “「ミトコンドリアを失った生物の軌跡」
~大規模解析で探るミトコンドリアの退縮~”. 筑波大学生物学類. 2025年5月25日閲覧。
^ John P.; Whatley FR. (1975). “Paracoccus denitrificans and the evolutionary origin of the mitochondrion.”. Nature 254 (5500): 495-498. PMID 235742.
^ a b Yang D.; Oyaizu Y.; Oyaizu H.; Olsen GJ.; Woese CR. (1985). “Mitochondrial origins” (pdf). Proc Natl Acad Sci 82 (13): 4443-4447. PMID 3892535.
^ a b Viale AM.; Arakaki AK. (1994). “The chaperone connection to the origins of the eukaryotic organelles.”. FEBS Lett. 341 (2-3): 146-151. PMID 7907991.
^ Andersson SG.; Zomorodipour A.; Andersson JO.; Sicheritz-Ponten T.; Alsmark UC.; Podowski RM.; Naslund AK.; Eriksson AS. et al. (1998). “The genome sequence of Rickettsia prowazekii and the origin of mitochondria.”. Nature 396 (6707): 133-140. PMID 9823893.
^ Rappe et al. (2002). “Cultivation of the ubiquitous SAR11 marine bacterioplankton clade”. Nature 418 (6898): 630-633. PMID 12167859.
^ Williams KP.; Sobral BW.; Dickerman AW. (2007). “A robust species tree for the alphaproteobacteria.” (pdf). J Bacteriol. 189 (13): 4578-4586. PMID 17483224.
^ 小野 興作; 大島 福造; 渡辺 漸 (1958). “Warburgの「癌細胞の起原」に就いて”. 岡山医学会雑誌 70 (12supplement): 143-154. doi:10.4044/joma1947.70.12supplement_143.
^ a b Hayashi, J., et. al. (2008). “ROS-generating mitochondrial DNA mutations can regulate tumor cell metastasis”. Science 320 (5876): 661-664. PMID 18388260.
^ NAKAMURA M.; OHSAWA S.; IGAKI T. (2014-10-17). “Mitochondrial defects trigger proliferation of neighbouring cells via a senescence-associated secretory phenotype in Drosophila (ショウジョウバエにおいて、ミトコンドリアの障害が、細胞老化関連分泌因子(SASP因子)を介して隣接細胞の増殖のトリガーになる)”. Nature Communications 5. doi:10.1038/ncomms6264 2020年1月5日閲覧。.
^ “井垣達吏教授らの研究成果が、英国科学誌「Nature Communications」に掲載されました。”. 京都大学 大学院 生命科学研究科 (2014年10月28日). 2020年1月5日閲覧。
^ “細胞間の相互作用で良性腫瘍ががん化する仕組みを解明” (html). 科学技術振興機構, 神戸大学 (2012年10月1日). 2020年1月5日閲覧。
関連項目
ウィキペディアの姉妹プロジェクトで
「ミトコンドリア」に関する情報が検索できます。
ウィクショナリーの辞書項目
コモンズのメディア
ウィキソースの原文
ウィキブックスの教科書や解説書
ミトコンドリアのシャトル系
葉緑体 – ミトコンドリアと同様に、外膜と内膜、さらに、独自のDNAも有する。
カルニチン
ハプログループF (mtDNA)
母の呪い(英語版)
オンコサイト(英語版)
表話編歴
細胞小器官
典拠管理データベース ウィキデータを編集
カテゴリ: 細胞小器官ミトコンドリア細胞呼吸
最終更新 2025年6月14日 (土) 17:18 (日時は個人設定で未設定ならばUTC)。
テキストはクリエイティブ・コモンズ 表示-継承ライセンスのもとで利用できます。追加の条件が適用される場合があります。詳細については利用規約を参照してください。』 -
物理学の要用
https://www.aozora.gr.jp/cards/000296/files/46686_25533.html『物理学の要用 福沢諭吉
物理学とは、天然の原則にもとづき、物の性質を明らかにし、その働を察し、これを採ってもって人事の用に供するの学にして、おのずから他の学問に異なるところのものあり。
たとえば今、経済学といい、商売学といい、等しく学の名あれども、今日の有様にては、経済商売の如き、未だまったく天然の原則によるものに非ず。
いかんとなれば、経済商売に、自由の主義あり、保護の主義あり。そのもとづくところ、同じからずして、英国の学者が自由をもって理なりといえば、亜国の人は保護をもって道みちなりといい、これを聞けば双方ともに道理あるが如し。
されば、経済商売の道理は、英亜両国においてその趣おもむきを異にするものといわざるをえず。
物理はすなわち然らず。
開闢かいびゃくの初より今日にいたるまで、世界古今、正まさしく同一様にして変違あることなし。
神代の水も華氏の寒暖計二百十二度の熱に逢うて沸騰し、明治年間の水もまた、これに同じ。
西洋の蒸気も東洋の蒸気も、その膨脹の力は異ならず。
亜米利加の人がモルヒネを多量に服して死すれば、日本人もまた、これを服して死すべし。
これを物理の原則といい、この原則を究めて利用する、これを物理学という。
人間万事この理に洩もるるものあるべからず。
もしあるいは然しからざるに似たる者は、未いまだ究理の不行届ふゆきとどきなるものと知るべし。
そもそもこの物理学の敵にして、その発達を妨ぐるものは、人民の惑溺わくできにして、たとえば陰陽五行論いんようごぎょうろんの如き、これなれども、幸にして我が国の上等社会には、その惑溺はなはだ少なし。
拙著『時事小言』の第四編にいわく、
「(前略)ひっきょう、支那人がその国の広大なるを自負して他を蔑視べっしし、かつ数千年来、陰陽五行の妄説に惑溺して、事物の真理原則を求むるの鍵を放擲したるの罪なり。
天文をうかがって吉兆を卜ぼくし、星宿の変をみて禍福を憂喜し、竜といい、麒麟きりんといい、鳳鳥ほうちょう、河図かと、幽鬼、神霊の説は、現に今日も、かの上等社会中に行われて、これを疑う者、はなはだ稀まれなるが如し。
いずれも皆、真理原則の敵にして、この勁敵けいてきのあらん限りは、改進文明の元素は、この国に入るべからざるなり。
我が日本にもこの敵なきに非ざりしかども、偶然の事情によりて大いに趣おもむきを異にするところあり。
我が国において、鬼神幽冥の妄説は、多くは仏者の預るところとなりて、もっぱら社会に流行したることなれども、三百年来、儒者の道、ようやく盛にして、仏者に抗し、これに抗するの余りに、しきりに幽冥の説を駁ばくして、ついには自家固有の陰陽五行論をも喋々ちょうちょうするを忌いむにいたれり。
たとえば、儒者が易経えききょうを講ずれども、ただその論理を講ずるのみにして、卜筮ぼくぜいを弄もてあそぶを恥ずるが如し。
その仏を駁撃するはあたかも儒者流の私わたくしなれども、この私論しろんの結果をもって惑溺を脱したるは、偶然の幸というべし。
支那の儒者も孔孟の道を尊び、日本の儒者も孔孟の書を読み、双方ともにその教の源みなもとを同じゅうして、その社会に分布したる結果において、まったく相反するは、偶然に非ずして何ぞや。
けだし、支那の儒教は敵なきがゆえに、その惑溺をたくましゅうし、日本の儒教は勁敵に敵して自から警いましめたるものなり。
かつ我が儒者はたいがい皆、武人の家に生れたる者にして、文采風流の中におのずから快活の精神を存し、よく子弟を教育してその気風を養い、全国士族以上の者は皆これに靡なびかざるはなし。
改進の用意十分に熟したるものというべし。」云々。
右の如く、我が国上等社会の人は、無稽の幽冥説ゆうめいせつに惑溺すること、はなはだ少なしといえども、その、これに惑溺せざるは、ただ一時仏者に敵するの熱心に乗じたるものにして、天然の真理原則を推究したる知識の働はたらきに非ざるがゆえに、幽冥説に向って淡白なるほどに、物理においてもまた自から漠然たるの情あるが如し。
儒者が地獄極楽の仏説を証拠なきものなりとて排撃しながら、自家においては、数百年のその間、降雨の一理をだに推究したる者なし。
雨は天より降るといい、あるいは雲凝こりて雨となるというのみにして、蒸発の理と数とにいたりては、かつてその証拠を求むるを知らざりしなり。
朝夕ちょうせき水を用いてその剛軟を論じながら、その水は何物の集まりて形をなしたるものか、その水中に何物を混じ何物を除けば剛水ごうすいとなり、また軟水なんすいとなるかの証拠を求めず、重炭酸加爾幾カルキは水に混合してその性を剛ならしめ、鉄瓶等の裏面に附着する水垢みずあかと称するものは、たいてい皆この加爾幾なりとの理は、これを度外におきて推究したる者あるを聞かず。
今日にありても儒者の教に養育しられたる者は、これらの談を聴きて瑣末さまつの事なりと思うべけれども、決して然らず。
欧州近時の文明は皆、この物理学より出でざるはなし。
彼の発明の蒸汽船車なり、鉄砲軍器なり、また電信瓦斯ガスなり、働の成跡は大なりといえども、そのはじめは錙朱ししゅの理を推究分離して、ついにもって人事に施したる者のみ。
その大を見て驚くなかれ、その小を見て等閑とうかんに附するなかれ。大小の物、皆みな偶然に非ざるなり。
人にして物理に暗く、ただ文明の物を用いてその物の性質を知らざるは、かの馬が飼料を喰くろうて、その品の性質を知らず、ただその口に旨きものはこれを取りて、然らざるものはこれを捨つるに異ならず。
然りといえども、馬はなお、その物の毒性なるか良性なるかを弁ずるの能力を有す。
然るに今の世の不学の徒は、汽車に乗りて汽の理をしらず、電信を用いて電気の性質を知らず、はなはだしきは自身の何物たるを知らずして、摂生の法を誤る者あり。
なおはなはだしきは、医は意なりと公言して、医術は憶測に出ずるものかと誤まり認したため、無稽むけいの漢薬を服して自得する者あり。
その愚の極度にいたりては、売薬をなめて万病を医せんと欲する者あり。
上等社会にしてその知識の卑しきこと、実に驚くに堪えたり。
ひっきょう物理を度外視するの罪にして、あるいは人にして馬に若しかずと評せらるるも、これに答うるの辞なかるべし。
我が慶応義塾において初学を導くにもっぱら物理学をもってして、あたかも諸課の予備となすも、けだしこれがためなり。
なお、その教則の事については他日陳述するところのものあるべし。
底本:「福沢諭吉教育論集」岩波文庫、岩波書店
1991(平成3)年3月18日第1刷発行底本の親本:「福沢諭吉選集 第3巻」岩波書店
1980(昭和55)年12月18日第1刷発行初出:「時事新報」時事新報社
1882(明治15)年3月22日発行入力:田中哲郎
校正:noriko saito
2006年12月30日作成青空文庫作成ファイル:
このファイルは、インターネットの図書館、青空文庫(http://www.aozora.gr.jp/)で作られました。入力、校正、制作にあたったのは、ボランティアの皆さんです。
』 -
福沢諭吉と物理学
https://webmagazine.ouj.ac.jp/matsui-tetsuo202404/『【教員コラム】福沢諭吉と物理学
松井 哲男
特任教授(自然と環境コース)皆さんは福沢諭吉と聞くと何を思い浮かべますか?明治日本の有名な教養人、『学問のすすめ』の著者、慶應義塾大学の創設者、(旧)1万円札の肖像、等々。およそこの表題にある物理とは全く縁のない人、と思っていませんか?
実は、彼が若い時に一番やりたかったのは物理学の研究だったのです。このことはあまり知られていないのですが、ここでそれを紹介したいと思います。
彼の口述筆記による『福翁自伝』によると、幕末の二十歳になったばかりの頃、大坂の緒方洪庵の「適塾」で学んでいたときに、蘭語で書かれた物理学書を手に入れてファラデーの電気説のこと等を知り、物理に非常に興味を持ったと書かれています。
彼は中津藩(大分県中津市)の下級藩士の出で、若い頃は儒学を学びましたが、だんだんそれに興味を失い西洋文明に傾倒していきます。
その橋渡しをしたのが物理学だったようです。
当時の日本は、「黒船ショック」で開国か攘夷かで騒然としていましたが、福沢は藩命で蘭学塾の講師として江戸に向かいます。福沢にとって物理学は、西洋医学と同様、西洋文明の優位性の背後にある「実学」だったようです。
蘭語を学んだのちも、独学で英語を学び、幕府がオランダから買い入れた咸臨丸に船長の木村摂津守の従者として乗り込んで、訪米しました。1860年のことです。
その後、幕臣の通訳として欧州諸国を1年程周り、またもう一度訪米して、海外で見聞したことを『西洋事情』などで紹介して、日本の西洋化の寵児となりました。
彼を明治新政府の役人として迎えたいという誘いもあったようですが、それはきっぱりと断り、彼は在野で日本の近代化に尽くしました。
彼は学問奨励の道を登りつめ、今日の日本学士院の前身である東京学士会院の初代会長を半年間務めています。
福沢が還暦を前にした1893年に三田の慶應義塾で行なった講演の速記録が残っています。
その題目は『人生の楽事』となっています。ここで「楽事」というのは、誰もが持つ趣味や志(こころざし)のようなもので、旅行好きな人、逆に家に一人籠っていたい人、絵画や骨董品を蒐集したい人、お金集めが好きな人、立身出世したい人など、千差万別あります。
福沢にとっての「楽事」とは物理学を学ぶことであり、本当は若い時から物理の研究をしたかったのだけれど、時代がそうさせてくれなかった、と告白しています。
そして、日本に物理学の小さい研究所を作りたかったとも言っており、具体的にどのくらいの費用が必要か現実的な試算もしています。
結局それもかないませんでしたが、ここで彼がこういう話をしたことを誰かが覚えていて、彼の死後にそれが実現されれば、草葉の影で感涙しているだろう、と話を締めくくっています。
福沢はこの講演の前年には北里柴三郎の伝染病研究所の設立に尽力したと伝えられていますが、彼が本当にやりたかったのは、自分が好きだった物理学の研究所を作ることだったようです。
(この記事は、2023年に執筆されました。)
松井 哲男 特任教授のプロフィールはこちら
公開日 2024-04-30 最終更新日 2024-07-03 』
-
HDD技術の変遷。MRから始まりHAMR/MAMR、そしてBPMまで
https://pc.watch.impress.co.jp/docs/topic/feature/2013099.html








『大原 雄介2025年8月1日 06:26
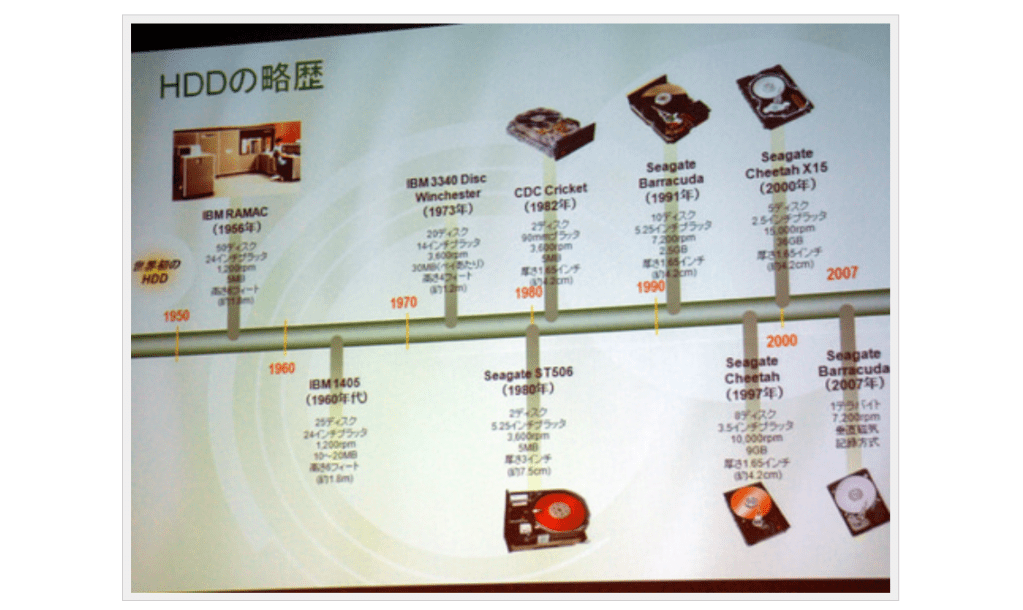
昨今のHDDに使われている技術の変遷を簡単にまとめてほしい、というお題をいただいた。ちなみにIBMのRAMACまで遡る必要はないと釘を刺されており、比較的最近(?)の話からのスタートとなる。
下敷きとなる話として、2008年にSeagateが都内で行なったワークショップの記事があったので、これをベースにしたい。つまり2008年までの話はそちらの記事を参考にしていただければ、という話である。
記事目次
現在のHDDの状況
MRやGMRといった磁気ヘッドの種類
現在研究中のBPM(Bit-Patterned Media)とは?
現在のHDDの状況実のところ、HDDの基本的な要素に関しては2000年前半にほぼ完成していると言ってよい。ステッピングモーターを利用してプラッタを駆動し、このプラッタの両面に磁気ヘッドを這わす形にして読み書きを行なうというものだ。プラッタの枚数は多くて11枚程度であり、HDDの容量はこのプラッタに保持できる記憶容量で決まる格好である。
以前はプラッタの回転速度を10,000rpmとか15,000rpmとかにすることでアクセス時間を減らす工夫がされていた。15,000rpmのものについては、3.5インチのままだと遠心力が強すぎてプラッタが壊れかねないというので2.5インチプラッタを採用した製品もあり、容量がそのぶん小さくなるという弊害すらあった。
2004年に登場した15,000rpmの3.5インチHDD「Ultrastar 15K147シリーズ」。容量は36GB、73GB、147GBだった
しかしこのようなものはもう存在しない。そもそも超高回転HDDは大規模なRAIDを組むことを前提にしたものだったが、こうした用途はエンタープライズSSDに置き換えがほぼ済んでしまっている。SSDのアクセス速度はもとより、容量としてもU.2で61TBの製品はすでに出荷されており、122TBの製品のサンプル出荷もスタートしている。
他方HDDは?というと最大でも30TB台でしかない。ただ容量価格比ではHDDはエンタープライズSSDの追従を許さない良好な数字を出しており、これもあって低頻度ながら定期的なアクセスがあるようなウォームストレージ向けに引き続き使われているという格好だ。
ウォームストレージ向けの場合、ランニングコストも重要になってくるため、強制冷却を掛けないと動作温度が上がりすぎてしまう10,000rpm以上のドライブはむしろ不適切であり、その結果、最近はエンタープライズ向けでも7,200rpmに抑えられている。こういう状況だから、転送速度のほうも推して知るべしである。プラッタ最外周をシーケンシャルアクセスするといった理想的な状況であれば300MB/s近い転送速度が期待できるが、そんな理想的な状況になることはまず考えられない。
であればインターフェイスはSATA 6Gbpsどころか3Gbpsでお釣りが来る計算になる。6Gbpsの定義がされたSerial ATA(SATA) Rev 3.0がリリースされたのは2008年だが、性能的には2004年のSATA Rev 2.0で十分間に合う形だ。SATAの仕様そのものは2021年のRevision 3.5aが最新となるが、HDDとの接続に関しては2008年のRevision 3.0でほぼ網羅した格好となっている。
SATAコネクタ
エンタープライズ向けにはSAS(Serial Attached SCSI)が使われることが多いが、こちらも途中からターゲットがHDDではなくSSDに切り替わりつつあり、SAS HDDに関しては2009年のSAS-2(6Gbps)ないし2013年のSAS-3(12Gbps)でほぼ網羅されており、2017年のSAS-4(22.5Gbps)に対応したHDDというのは見かけたことがない。実際転送速度で考えればSAS-3どころかSAS-2でもお釣りが来る計算だから、これは致し方ないだろう。
ほかの例としては、磁気ヘッドを搭載したアームを2組搭載することで、転送速度を倍増させられる技術もあって、たとえばSeagateは2017年12月にMulti Actuator Technologyを発表、2018年3月にはデモも行なわれたが、今のところ製品が出てくる話にはまったくなっていない。
Multi Actuator Technologyのイメージ
また2010年頃にはHDDにSSDをキャッシュ用に搭載するSSHD(Solid State Hybrid Drive)あるいはHybrid HDDと呼ばれる製品も登場したが、PC用のBoot DriveがほぼM.2 NVMe SSDに移行し、しかも十分な容量(~4TB)が提供される様になると、こうしたHybrid製品のニーズも次第に減ってきたようで、最近ではあまり見かけなくなった。
Seagateの2.5インチSSHD「Momentus XT」
MRやGMRといった磁気ヘッドの種類
そんなわけで、HDDに関する技術革新の中心は、ほぼ容量の増加に集中することになる。要するに、1枚のプラッタにどれだけのデータを記録できるかという競争である。
と言ってもプラッタの面積そのものは限界サイズまで大きくなっているから、後は記録密度を向上させるしか容量を増やす方法がないことになる。かくして、記録密度向上に向けての努力が積み重ねられることになる。
記録密度の第1の方法が、磁気ヘッドの改良である。HDDに利用される磁気ヘッドは1990年以降、次の技術が使われてきた。
1990年 : MR/AMR(Magneto Resistance/Anisotropic Magneto Resistance)
2000年 : GMR(Giant Magneto Resistance)
2004年 : TMR(Tunnel Magneto Resistance)
2024年 : HAMR/MAMR(Heat Assisted Magnetic Recording/Microwave Assisted Magnetic Recording)そもそもHDDは、以下の形で動作を行なう。
読み出し(リード)
記録媒体(つまりプラッタ)から磁気の方向を読み出して、それをデータとして出力
書き込み(ライト)
データに合わせて記録媒体に磁気の方向を変更して書き込み(上書き)さて、記録密度を上げるということは、1bit分の記録を行なう面積がそれだけ減るということになる。これは書き込みを行なう際にも問題であるが、それよりも読み込みの際に「読み取れる磁荷の量」が減ってしまうという問題が生じることになるのが大問題である。
MR(Magneto Resistance)
これを解決するための方法として最初に採用されたのがMRである。Magneto Resistance(磁気抵抗)という名前の通り、磁場(厳密に言えば磁束密度)に応じて抵抗値が変わるという効果を利用したものだ。
この効果、最初に発見されたのは1856年と古いのだが、当時はまだ材質の問題があり、抵抗値の変化が少なかったこともあって、あまり利用されなかった。ところがその後、InSb(インジウム・アンチモン)などの新材料の発見などもあり、従来の方法(磁場の誘導電流を利用する方法)よりも高い感度で磁場の測定ができるということで、1990年頃からMR素子を磁気ヘッドに利用するようになった。
ちなみに続くGMRとかTMRも括りとしてはMRに入るということで、あえてAMRと呼ぶ場合もある。初期のMRセンサーは、異方性磁気抵抗効果(Anisotropic Magneto Resistance)と呼ばれる性質をそのまま利用したもので、センサーとなる強磁性体と磁場の向きが一致している場合と一致していない場合で、抵抗値が概ね3~5%ほど異なるというもので、抵抗値を測定してこの差を読み取る仕組みとなっていた。
GMR(Giant Magneto Resistance)
続いて2000年頃に採用が始まったのがGMR(Giant Magneto Resistance)である。これは2つの強磁性体の間に非強磁性金属を挟み込むことで、抵抗値の変動がより大きくなる(2~3倍)という特徴を生かしたものだ。抵抗値の変動が大きい(≒感度が高い)というのは、わずかな磁場の変化でも捉えやすくなるということでもあり、これによりプラッタ上の記録面積を削減しても、AMRの場合と同等のレベルでの検出が可能ということになる。
TMR(Tunnel Magneto Resistance)
そのGMRに続いて2004年頃から実用化されたのがTMR(Tunnel Magneto Resistance)である。構造的にはGMR同様に2つの強磁性体の間に非強磁性金属を挟み込む構造だが、電流の掛け方がGMRと異なる方式である。こちらはトンネル効果を利用したものだが、抵抗値の変化率がAMR比で30倍以上と極めて大きいのが特徴であり、これによりGMRと比較しても大幅に感度を上げられることになった。
ということは、それだけ記憶領域の面積を小さくできる≒高密度化が達成できることになる。実際2004年からTMRを利用した磁気ヘッドが登場し始め、そこから20年を経過した現在も普及帯の製品はこのTMR磁気ヘッドが採用されている。今後も小容量なHDD向けには引き続きTMR磁気ヘッドが利用されることになるかと思う。
PMR(Perpendicular Magnetic Recording)
次のHAMR/MAMRについては後で説明することにして、先にヘッド以外の改良技術についても説明しておきたい。まずTMRとほぼ同時期に登場したのがPMR(Perpendicular Magnetic Recording : 垂直記録方式)である。このPMR、2005年にHGST(日立グローバルストレージテクノロジーズ)が行なった記者発表会の様子がこちらの記事に纏まっている。
それまでは面内記録方式あるいは長手記録方式と呼ばれる、つまりプラッタに円周状に設けられたトラックに沿う形で記録していたのだが、垂直記録方式ではこれをプラッタに対して垂直に記録するようにしている。それだけ高密度にできるというわけだ。
PMRを最初に商品化したのは東芝であるが、当初は3.5インチ未満の小型HDD(東芝は当初1.8インチサイズに採用した)の記憶容量増加が主目的であった。ただしすぐに3.5インチHDDにも採用されるようになっている。PMRでは、プラッタ上に占める面積は非常に小さいことになるので、当然ながらTMR磁気ヘッドとの併用で実用化した格好だ。
東芝が開発したPMR方式の1.8インチHDD
このPMRの性能を強化したのがWDのePMR(Enhanced Perpendicular Magnetic Recording)と呼ばれる技法である。これは書き込み時に少し大きめの電流を流すことで書き込み信号が安定し、結果的により記録密度を引き上げられる、というものである。
2023年にはこれを採用した28TB品のサンプル出荷が開始されることが報じられ、2024年10月には最大32TB品の出荷が開始された。
ePMRを採用した「Ultrastar DC HC690 SMR HDD」
ヘリウム充填
次に採用された技術がヘリウム充填である。そもそもHDDの中は真空ではなく(真空だとヘッドがプラッタに張り付いてしまう危険性がある)空気が密閉されている。もちろんごみなどが混入するとクラッシュの要因になるので、ゴミのない綺麗な空気を密閉しているわけだが、記録密度の向上に伴ってヘッドの位置制御が非常に困難になってきた。
ヘリウムガスが充填されたSeagateの「Exos X14」
空気分子の大半はN2(窒素)で分子量28.0134g/mol、次がO2(酸素)で分子量31.999g/molであるが、7,200rpmでプラッタが回転すると、これに伴いHDD内部の空気も結構な速度で動くことになる。
この空気がHDDのヘッドを支えるアームにぶち当たることで、アームがある程度振動することは避けられない。フラッターと呼ばれるこの現象を完全に抑えるのは非常に難しいので、ある程度許容せざるを得ないのだが、そうなると多少フラッターによってアームが上下してもヘッドがプラッタに接触しないようにマージンを取る必要がある。これが理由で、1台のHDDに搭載できるプラッタの枚数は5枚かその位に制限されていた。
ところがここでHDDの内部にHe(ヘリウム)を充填するというアイデアが生まれた。理由は簡単で、分子量がはるかに小さい(4.003g/mol)ことだ。つまり同じ量のN2やO2分子に比べると、ずっと軽い。軽いということは、それだけアームにフラッターを起こさせる力が小さいということで、この結果接触防止のために必要なマージンとして取るべき高さ方向の幅を大幅に減らすことが可能になった。
この結果、1台のHDDに10枚以上のプラッタを収められるようになる。プラッタの枚数はHDDの容量にそのまま直結するから、このヘリウム充填は容量増加に大きく貢献した。
ちなみにH2(水素)にすればさらに分子量は減る(2.016g/mol)し、ヘリウムに比べるとはるかに安価であるが、以下のような問題があるので、採用にはいたっていない。
ヘリウムを水素にしても、それほど大きく枚数が増やせるわけではないというか、ヘリウムを利用した時点でもう限界まで枚数を増やしており、これ以上枚数を増やすのはかなり難しい。
水素は密閉が非常に困難でさっさと抜けてしまうため、本気で密閉しようとすると現在の3.5インチHDDの大きさではかなり困難が伴う。ヘリウムは従来の密閉技術と同じとまでは言わない(実際IBMが2015年にヘリウム充填HDDを発売するにあたっては、内部構造とか充填技法の確立にかなり苦労した)までも、3.5インチHDDの大きさに収めることが技術的に可能である。
水素が漏れてしまい、酸素と結合すると爆発の危険性がある。ヘリウムは不活性ガスなのでこうした危険性がない。
SMR(Shingled Magnetic Recording)
もう一つ忘れてはいけないのが瓦書きことSMR(Shingled Magnetic Recording)である。SMRの技術的な構造はこちらの記事に纏まっているのでご覧いただきたい。ただSMRはシーケンシャルアクセスには適しているが、ランダムアクセス時の性能は悪化するというレベルではないほどに劣化する。ちなみに従来型の記録方式はCMR(Conventional Magnetic Recording)と呼ばれる。
SMR方式では、プラッタの外周にCMR方式の一時記憶領域(これをMedia Cacheと呼ぶ)を確保し、ランダムな書き込みはまずこのMedia Cacheに書き込み、後でSMR方式の領域に書き戻すというWrite Back方式を採用するが、Media Cacheの容量にも限りがあるし、SMRは原理的に先頭からシーケンシャルに書き込む必要があるので、ひたすら書き込むだけという用途(たとえば監視カメラの記録用ストレージ)には適しているが、一般的な利用に適しているか?と言われるとちょっと疑問ではある。
またSMRを採用して増える容量は10%とか20%のオーダーであって、2倍とか3倍には遠く及ばない。このあたりもあってか、HDDメーカーは最近では明確にSMR方式かCMR方式かを明示している。
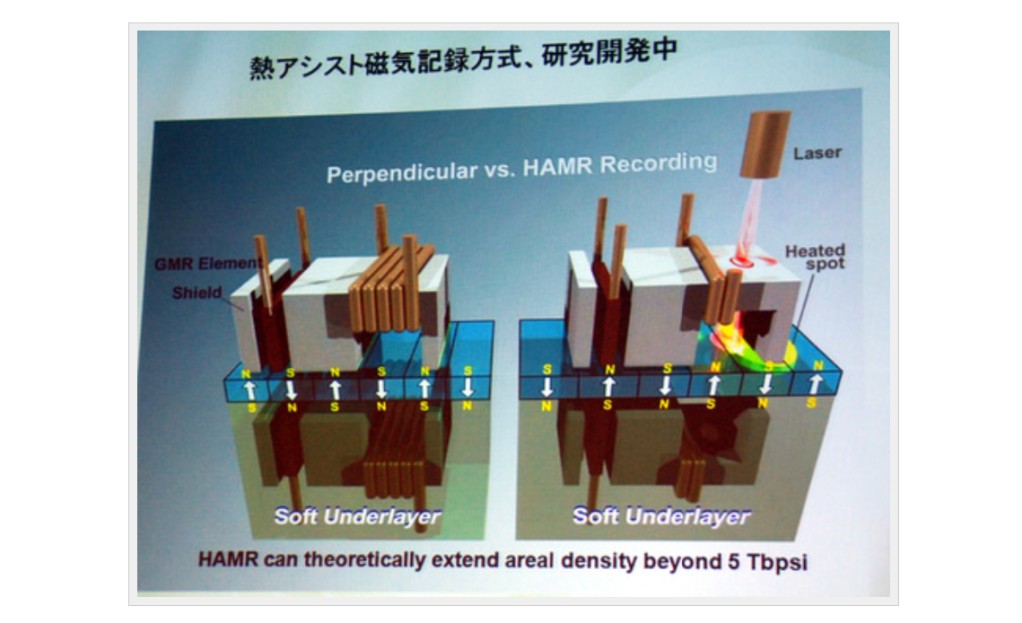
HAMR(Heat-Assisted Magnetic Recording)
MAMR(Microwave-Assisted Magnetic Recording)
ということで最後に昨今のホットトピックであるHAMR(Heat-Assisted Magnetic Recording)と、MAMR(Microwave-Assisted Magnetic Recording)について紹介したい。
まとめて熱アシストと呼ばれるHAMR/MAMRは再びヘッド側の技術ではあるのだが、これまでのAMR→GMR→TMRが読み取り性能の向上だったのに対し、HAMR/MAMRは書き込み能力の向上のための技術である。
背景にあるのは、TMR+PMRの組み合わせによる記録密度向上に行き詰まりが出て来たことだ。実を言えば、TMR方式そのもので言えば、まだ記録密度向上自体は可能である。つまり読み出しだけで言えば、さらに記録密度が上がっても対応できる。ところが書き込みのほうが追い付かなくなった格好だ。
要するに、記録面積を小さくしても十分な磁束密度を確保すると同時に、熱揺らぎ(温度上昇に伴い、磁気が消えたり反転したりする現象)への対策として、プラッタに高保磁力材料を利用したところ、今度は書き込みが簡単にできなくなってしまった。
この対策として考え出されたのが、「書き込み時のみ熱を加える」という仕組みだ。熱揺らぎは要するに温度が上がると磁気の状態が変化しやすいということであり、これを逆手に取って書き込む瞬間のみ温度を上げてやれば簡単に書き込みが行なえる。
終わった瞬間に加熱をやめればすぐに常温に戻り、書きこんだ状態が堅固に保持される、というわけだ。この「書き込む時に加熱する」という仕組みから、熱アシストと呼ばれるわけだ。この熱アシストにより、記録密度をTMR+PMRと比べて将来的には3倍以上向上できる、とされている。
仕組みは簡単であるが、実装するのは決して楽ではない。実際冒頭で紹介した2008年のSeagateのワークショップでも、この熱アシストについて言及がある。
実際のところ、TMR磁気ヘッドの量産が始まった2004年頃から、すでに熱アシスト方式の研究が始まっており、ただし量産までに20年かかったという代物である。この熱アシスト、大別すると以下のの2種類がある。
レーザーを当てて加熱する方式(HAMR)
マイクロ波を当てて加熱する方式(MAMR)
このどちらが良いのか?というと一長一短というか、どちらの方法にも技術的に困難な課題があり、それを克服するのに時間を要した結果が20年という長きに渡ってTMR+PMRが使われ続けた理由である。
HAMRの方は実装は容易だが、ヘッドも一緒に加熱されてしまう。結果的に、MAMR用のヘッドに比べると寿命が1桁短いなんていう話もあったりする(SeagateはMAMR用ヘッドと同等の寿命を確保したとしている)。
一方でMAMRは?というとまだ現時点でHAMRほどの性能が出ていない。MAMR型で先行しているのは東芝で、元々同社は2021年にFC-MAMR(Flux Control-Microwave Assisted Magnetic Recording: 磁束制御型マイクロ波アシスト磁気記録方式)と呼ばれる方式を利用した18TBのHDDのサンプル出荷を開始している。
ただ、この時の構成はプラッタ9枚で16TB→18TB、つまりプラッタあたりの記憶容量増加は12.5%と非常に限られた記憶容量増加に過ぎない。MAMR方式の本命はMAS-MAMR(Microwave Assisted Switching Microwave Assisted Magnetic Recording 共鳴型マイクロ波アシスト磁気記録方式)と呼ばれる方式を予定しているが、こちらは量産に入るまでまだ時間がかかると見られている。もともとFC-MAMR方式はMAS-MAMR方式の開発の中で生まれた技術である。まずは手堅い技術で実績を積もう、というわけだ。
Seagate
ちなみに実際のところで言うと、SeagateはHAMRを採用。2025年1月に、HAMRベースとなるMozaic 3+を搭載したExos M HDDのサンプル出荷を開始している。プラッタ容量は3.6TBで、10枚構成で最大36TBの容量とされる。
東芝
東芝はHAMRとMAMRの両対応を想定している。先に書いたように、すでにFC-MAMR方式の製品はすでに出荷しているが、STO-MAMR方式の方はまだ時間がかかる。
そこで先行するSeagateに負けじと、まずはHAMR方式の製品を2025年中に最大32TB品をサンプル出荷開始予定である。長期的にはMAS-MAMR方式に切り替えてゆくとするが、この切り替えには数年かかると同社は見ている。
Western Digital
2017年にはMAMR方式の優位性を説いたWDだが、2025年2月に開催されたInvestor Day 2025におけるロードマップを見ると、今年いっぱいは既存のePMRのまま最大36TB程度まで容量を増やし、2026年度にHAMRを利用する予定であると示されている。
そんなわけで2025年で言えば各社HAMR+PMRという格好で30TB超えを達成し、このまま40~50TB位まで容量を増やせることを想定している。途中でHAMR+PMRがMAMR+PMRに切り替わるかもしれないが、これはMAMRの技術開発の進捗次第である。
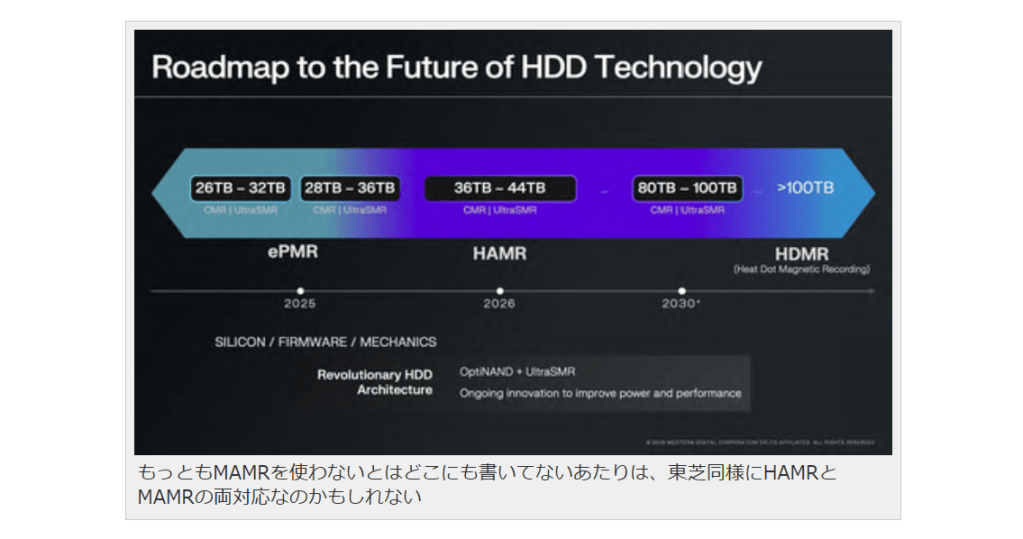
現在研究中のBPM(Bit-Patterned Media)とは?
ではその先は?という話を最後にご紹介しておくと、現在研究されているのがPatterned MediaあるいはBPM(Bit-Patterned Media)と呼ばれるものである。以下の画像で2030年以降にHDMR(Heat Dot Magnetic Recording)と書かれているのがそれだ。
もっともMAMRを使わないとはどこにも書いてないあたりは、東芝同様にHAMRとMAMRの両対応なのかもしれない
現在のプラッタは、ガラスあるいはアルミニウムの円盤の上に磁性体の被膜を蒸着させる形で構成されている。なので複数の磁性体粒子で1bit分の記憶領域を構成する形だが、記録密度を上げてゆくと隣接する粒子同士の相互干渉により安定状態を保つのが難しくなる。
そこで1bit分の記録を行なう1つの磁性体粒子を人工的に並べてやることで、相互干渉を防ぎながらもっと記録密度を上げられるというのがPatterned Mediaの基本的な原理である。実はこれも昔から研究されている事柄であり、TMRC 2010(The 21st Magnetic Recording Conference)で東芝がBPMを試作し、2.5Tbit/平方インチの記録密度を達成したことを発表している。
もっともこの時はBPMの試作に成功しただけで、これを自由に読み書きすることはできなかったのだが……。ちなみにこの試作したBPMは、磁性体粒子の直径が17nmだったそうだ。
現時点でもまだ実用に耐えるBPMは完成していない。ロードマップでHDMRの登場時期が明確にされていないのは、いつ実用に耐えるレベルのBPMが完成するかはっきりしないためである。とりあえずはそこまでの間、HAMRないしMAMRにPMRを組み合わせる形でなんとか容量増加に向けて取り組んでゆく、という格好になるかと思われる。』